Anmerk
Start Weiter
7
Eine
endlose Zahl der schönsten Formen Leakey 1995
121-151
Der
letzte Satz von Darwins
<Entstehung der Arten> ist so berühmt wie lyrisch:
»Es
liegt etwas Großartiges in dieser Ansicht vom Leben, wonach es mit allen
seinen verschiedenen Kräften aus wenig Formen, oder vielleicht nur einer,
ursprünglich erschaffen wurde; und daß, während dieser Planet gemäß
den bestimmten Gesetzen der Schwerkraft im Kreise sich bewegt, aus einem
so schlichten Anfang eine endlose Zahl der schönsten und wundervollsten
Formen entwickelt wurden.«1
Als
vor etwa fünf Millionen Jahren die erste Menschenspezies entstand, war sie
eine jener »schönsten Formen«, und das gleiche gilt heute für uns,
den Homo sapiens. Als Produkte der Wechselfälle des Lebens, entstanden
durch das Wechselspiel der Evolutionsprozesse und der manchmal launischen Hand
des Aussterbens, bilden wir und die anderen Arten, mit denen wir die Erde
teilen, eine Lebensgemeinschaft von fast beispielloser Vielfalt.
Wie
wir im vorangegangenen Kapitel gesehen haben, hat der Motor der
Evolution in den letzten 100 Millionen Jahren insgesamt für eine Zunahme der
Artenvielfalt gesorgt, die in der Geschichte der vielzelligen Lebensformen
nicht ihresgleichen hat, abgesehen von ihren explosionsartigen Anfangen. Kurz
und auffällig unterbrochen durch das Aussterben am Ende der Kreidezeit,
führte diese Zunahme der Vielfalt bis heute zu einer Welt, in der mehr Arten
leben als in jeder anderen Phase der Erdgeschichte.
Für
den Paläontologen wird die Reichhaltigkeit des Lebendigen in den Fossilfunden
auf augenfällige Weise deutlich.
Im
Mittelpunkt meines Berufslebens stand zwar über viele Jahre hinweg die
Geschichte der Menschenfamilie, aber ich war mir immer sehr deutlich des
größeren Zusammenhanges bewußt, in dem sie sich entfaltete. Die
Lebensräume Ostafrikas, die mir am vertrautesten sind, haben in den letzten
15 Millionen Jahren starke Wandlungen durchgemacht, die in erheblichem Ausmaß
auf geologische Veränderungen zurückgingen.
Betrachtet
man diese 15 Millionen Jahre als großes Schauspiel, dann gab es in
wechselnden Szenen komplexer Lebensgemeinschaften eine ständige Veränderung
der Rollenbesetzung. Jede dieser Lebensgemeinschaften war für sich
gesehen vollständig, aber wie man an den Fossilfunden erkennt, hatte sie auch
immer eine Vergangenheit und natürlich eine Zukunft. Jede war ein
vorübergehender Ausdruck des Lebensflusses. Wie ich in Kürze erläutern
werde, ist die paläontologische Sichtweise für die derzeitige biologische
Vielfalt nach meiner Überzeugung von großer Bedeutung; es ist eine
Sichtweise, die erst seit kurzem ins Blickfeld der Ökologen rückt.
Als
ich die Leitung der Naturschutzbehörde übernahm, wurde ich aus der
Vergangenheit sehr plötzlich in die Gegenwart versetzt, von dem vorrangigen
Interesse für ausgestorbene Arten zu der Sorge um Arten, die vom Aussterben
bedroht waren. Die natürliche Tierwelt Kenias ist äußerst vielfältig und
steht dem Artenreichtum in fast allen anderen Gebieten der Welt in nichts
nach. Als Direktor hatte ich anfangs die Aufgabe, Notfallmaßnahmen zu
ergreifen und der verheerenden Wilderei insbesondere bei Elefanten und
Nashörnern ein Ende zu machen. Dabei blieb die umfassendere Artenvielfalt
meines Landes zwangsläufig außen vor — jedenfalls eine Zeitlang.
Aber
in den ruhigen Augenblicken des Nachdenkens, in denen ich den
drängenden Erfordernissen des Amtes entkam und beispielsweise in einem
kleinen Flugzeug von Nairobi südwärts zum Tsavo-Nationalpark und weiter nach
Westen nach Masai Mara an der Küste reiste, oder bei den allzu seltenen
Gelegenheiten, wenn ich nach Norden zum Turkanasee flog, wurde ich an den
Reichtum des Lebens unter mir erinnert.
122/123
In
der Ökologie kennt man für die Artenvielfalt oder Biodiversität dreierlei
Maßstäbe. Der erste, Alpha-Diversität genannt, ist ein Maß für die
Artenzahl in einer ökologischen Lebensgemeinschaft. Die Beta-Diversität
ergibt sich aus dem Vergleich der Artenzusammensetzung in benachbarten
Lebensgemeinschaften, die sich in bestimmten physikalischen Eigenschaften
(beispielsweise der Höhenlage) unterscheiden. Und die Gamma-Diversität, der
dritte Maßstab, sagt etwas über die Lebensgemeinschaften eines größeren
geographischen Gebietes aus und umfaßt unter Umständen Gegenden mit
ähnlichen Lebensräumen, die viele Kilometer voneinander getrennt sind. Der
Flug von Nairobi zum Turkanasee bietet Anschauungsunterricht für alle drei
Maßstäbe.
Nairobi
liegt etwa 1500 Meter hoch auf dem geologischen Dom, der die kontinentale
Kruste vor 15 Millionen Jahren von einer Höhe knapp über dem Meeresspiegel
bis fast auf 3000 Meter an der höchsten Stelle anhob. Wenn ich von Nairobi
aus nach Norden fliege, muß ich die Böschung des Rift-Tals überwinden, die
von der Stadt aus etwa 1200 Meter ansteigt. Es ist eine äußerst fruchtbare
Gegend: Auf dem nährstoffreichen, roten Vulkanboden breitet sich ein
Flickenteppich aus Tee- und Kaffeeplantagen aus, in den kleine Städte
eingestreut sind. Die Bewältigung des Talabhanges ist immer ein dramatisches
Ereignis: In alle Richtungen erstreckt sich interessantes Gelände, und man
hat gigantische Wolkenformationen über und unter sich. Ich bin jedesmal
erleichtert, wenn ich die gefährliche Kante hinter mir habe.
Im
Westen fallen die Talflanken jäh ab und schaffen so einen Kontrast zwischen
der grünen Hochebene und der ausgedörrten Talsohle. Die Aberdare-Berge im
Osten der Flugroute werden reichlich mit Feuchtigkeit versorgt und beherbergen
eine wunderbar vielfältige Tierwelt mit den eleganten schwarzweißen
Stummelaffen und sogar mit Leoparden. Früher lebten hier auch Zehntausende
von Elefanten, aber so viele sind es heute nicht mehr, denn ihre Zahl wurde
von Wilderern und durch die Ausweitung der Landwirtschaft dezimiert; derzeit
leben hier noch etwa fünftausend von ihnen. Im Osten jenseits der
Aberdare-Berge liegt der Mount Kenya, dessen schneebedeckter Gipfel sich bis
über 5000 Meter erhebt.
124
Ein
einziger Ausblick bietet gewaltige Gegensätze: vergletscherte Gipfel,
Bergwiesen und dichte, kühlere Wälder am Mount Kenya; üppige Feuchtwälder
an den unteren Abhängen der Aberdares; trockene Wüste an der Talsohle; und
dazwischen ein kompliziertes, abgestuftes Vegetationsmosaik. Ich wußte, daß
es in jedem dieser Lebensräume mit Ausnahme der kalten Gletscher ein höchst
vielfältiges Tier- und Pflanzenleben gab, das heißt eine hohe
Alpha-Diversität. Und wenn man von den unteren zu den höheren Abhängen des
Mount Kenya aufstieg, traf man auf völlig unterschiedliche biologische Welten
vom Subtropischen bis zum Alpinen; die Beta-Diversität war also ebenfalls
hoch.
Auf
dem fast dreistündigen Flug von Nairobi zum Ostufer des Turkanasees zieht
unter mir eine atemberaubende ökologische Vielfalt vorüber, die alle
gerade beschriebenen Lebensgemeinschaften und noch vieles andere umfaßt. Ich
überquere die Kante der Laikipia-Hochebene, und danach, während der letzten
eineinhalb Flugstunden, eröffnet sich die Aussicht auf Lavaströme und
Krater, ausgetrocknete Seen, die Schatten versiegter Wasserläufe in der
trockenen Erde und schließlich die Sandsteinformationen, die das Ostufer des
riesigen Sees bilden. Wo wir landen, ist die Alpha-Diversität natürlich
geringer, bestimmt durch geringe Niederschlagsmengen und hohe Temperaturen.
Dennoch
gibt es in dieser Gegend mehr Leben, als die meisten Besucher auf den ersten
Blick bemerken; es reicht aus, damit hier Herden von Weißschwanzgnus und
Leierantilopen, Löwenrudel und sogar Leoparden leben können. Der Flug
vermittelt einen kleinen Eindruck von Kenias Gamma-Diversität, denn man kann
die Lebensgemeinschaften in einem recht großen geographischen Bereich
vergleichen; sie ist ebenfalls hoch, und man erkennt sofort, wie wichtig die
unterschiedliche Geländegestalt für die Entstehung dieser Vielfalt ist. Das
reichhaltige Mosaik der verschiedenen Höhenlagen mit seinen unzähligen
Lebensräumen, in denen die täglichen Temperaturschwankungen, die
Feuchtigkeit und viele Bedingungen des Mikroklimas immer wieder anders sind,
läßt ein ebenso reichhaltiges Mosaik ökologischer Gemeinschaften nicht nur
gedeihen, sondern bringt sie sogar hervor.
125
Würde
ich von Nairobi aus nicht nach Norden, sondern nach Westen fliegen, über das
Rift-Tal nach Uganda und noch weiter, so würde sich vor meinen Augen schon
bald ein undurchdringliches Grün ausbreiten, ein Teppich aus tropischem
Regenwald von Horizont zu Horizont. Er ist die Heimat der afrikanischen
Menschenaffen. Tropische Regenwälder beherbergen eine gewaltige biologische
Vielfalt, das stimmt, aber ohne die Geländeunterschiede des Rift-Tales fehlen
ihnen die Gegensätze, die ein wichtiges Element der Gamma-Diversität sind;
hier gibt es keine Herden von Steppentieren, keine Eidechsen, die an das Leben
in trockenen Wüsten angepaßt sind, keine Bergblumen.
Das reich gegliederte
Gelände im Osten des Rift-Tales mit seinem Mosaik der Lebensräume
lieferte neben vielem anderen auch den Impuls für die erste Evolution der
Menschenfamilie.
Ich
habe über meine Erfahrungen mit der Paläontologie und mit dem Naturschutz in
Kenia gesprochen, weil ich damit das Thema dieses Kapitels einführen will:
den Begriff der biologischen Vielfalt in der heutigen Welt. Jetzt möchte ich
den Blick über Kenia und über Afrika hinaus auf die ganze Welt lenken, und
dabei werde ich mich mit zwei Fragen beschäftigen, die beide für die
moderne, ökologisch orientierte Evolutionsforschung von zentraler Bedeutung
sind. Die erste betrifft die Form der biologischen Vielfalt und die Vorgänge,
durch die sie im lokalen und globalen Maßstab entsteht. Wie wir sehen werden,
kann man die Form mit einer gewissen Zuverlässigkeit beschreiben, denn sie
ist einfach so, wie man es deutlich vor Augen hat, aber die Vorgänge, die
ihrer Entstehung zugrunde liegen, sind nicht so offensichtlich. Die zweite
Frage lautet: Wie viele Arten bilden weltweit die biologische Vielfalt, zu der
auch wir gehören?
Sie zu beantworten mag einfach erscheinen, aber das stimmt
nicht. Ich werde erklären, warum.
Die
Gesamtform der biologischen Vielfalt wird von zahlreichen Faktoren bestimmt,
aber ich möchte mich auf die beiden wichtigsten konzentrieren. Der erste ist
die weltweite Verteilung der Lebensformen: Wo findet man die größte Zahl von
Arten? Der zweite hat mit dem Vergleich der biologischen Vielfalt in den
Meeren und an Land zu tun. Beide sind durch die Triebkraft der Evolution und
die Dynamik der Ökosysteme innig miteinander verknüpft.
126
Das
auffälligste Merkmal der biologischen Vielfalt auf der Erde ist die
ungleichmäßige Verteilung. Vereinfacht gesagt, ist die Artenvielfalt
beiderseits des Äquators am größten; mit höherer geographischer Breite,
das heißt in Richtung des Nord- und Südpols, wird sie immer geringer. Wer
beispielsweise in Nordamerika oder Europa zu Hause ist und dann einmal nach
Kenia kommt, bemerkt den Unterschied sofort. Er zeigt sich im ganzen Spektrum
der Tierwelt, nicht nur bei dem gewaltigen Anblick der wandernden Tierherden
und an den Löwen, Leoparden und Geparden, die den Besucher so stark
beeindrucken, sondern auch an dem phantastischen Reichtum der Vogelwelt und
natürlich am Gewimmel der Insekten. Ebenso deutlich wird das Prinzip an den
Pflanzen, insbesondere in den Regenwäldern mit ihrer reichhaltigen Mischung
der Baumarten und der auf ihnen wachsenden Epiphyten, die ihrerseits wieder
von Mikroepiphyten besiedelt sind. Auch wenn man sich auf die mikroskopische
Ebene begibt, zu den Bakterien und einzelligen Pilzen, trifft man auf den
gleichen Überreichtum der kenianischen Lebensformen, der den Artenreichtum in
den Heimatländern der meisten Besucher bei weitem übertrifft.
Diese
energische Handschrift der Natur, »breitengradabhängiger Gradient der
Artenvielfalt« genannt, ist den Biologen schon seit langem bekannt. Mit
zahllosen Hypothesen versuchte man ihn zu erklären, und das führte letztlich
zur Entstehung der theoretischen Ökologie. Er ist auch für den Artenschutz
von beträchtlicher Bedeutung: Wenn man in den Tropen einen Quadratkilometer
Lebensraum zerstört, gefährdet man damit mindestens zehnmal so viele Arten
wie beim Verlust der gleichen Fläche in gemäßigten Klimazonen. Besonders
reichhaltig ist die biologische Vielfalt in den tropischen Regenwäldern: Sie
bedecken ein Sechzehntel aller Landflächen der Erde, beherbergen aber über
die Hälfte aller Arten. Deshalb ist die erbarmungslose Zerstörung dieser
Wälder zutiefst beunruhigend.
Eine
endlose Zahl der schönsten Formen 127
Die
Konzentration auf die Regenwälder sollte aber, wie eine Studie kürzlich
gezeigt hat, nicht auf Kosten anderer Lebensräume gehen. Wie der Biologe
Michael Mares von der University of Oklahoma in einer Übersichtsuntersuchung
an fast 1000 südamerikanischen Arten feststellte, findet man die größte
Vielfalt zumindest bei manchen Artengruppen überraschenderweise in trockenen
Gegenden, beispielsweise in den Steppen (Llanos) Venezuelas und Kolumbiens,
den Gras- und Buschlandschaften (Cerrados und Caatingas) Brasiliens und den
argentinischen Pampas. »Trockengebiete gelten oft als Regionen mit geringer
biologischer Vielfalt«, stellt Mares in der Fachzeitschrift Science fest,
»aber was die Säugetiere angeht, sind sie die artenreichsten Gegenden des
Kontinents.«2)
Mares'
Entdeckung mindert den Wert der Regenwälder als Regionen großer biologischer
Vielfalt nicht, aber sie erweitert unsere Kenntnisse darüber, wo man Vielfalt
antreffen kann. In einem Kommentar zu dem Aufsatz von Mares schrieben die
Ökologen Stuart Pimm und John Gittleman von der University of Tennessee:
»Wir wissen zu wenig darüber, wo die Artenvielfalt ist, warum sie dort ist
und was aus ihr werden wird.«3)
Bevor
ich mich mit ein paar mutmaßlichen Gründen für den tropischen Artenreichtum
beschäftige, möchte ich einige kurze Beispiele für das Ausmaß der
Unterschiede anführen. Ein Ameisenspezialist, der von Alaska nach Brasilien
wandern und unterwegs in jeder Region die Arten zählen würde, träfe am
Ausgangspunkt der Reise auf drei und am Ziel auf 222 Ameisenarten. Das ist
ein Unterschied von fast zwei Zehnerpotenzen. Der beste Ameisenfachmann ist
natürlich der Biologe Edward Wilson von der Harvard University; er erklärte
1987 in Washington auf einer wissenschaftlichen Tagung über die
Artenvielfalt: »Ich habe an einem einzigen Baum in Peru 43 Ameisenarten
identifiziert, ungefähr ebenso viele wie auf den gesamten Britischen
Inseln.«
Das
gleiche würden Vogel- oder Baumliebhaber erleben. Peter Ashton zum Beispiel,
ein weiterer Biologe der Harvard University, zählte in Borneo die Baumarten
auf 100.000 Quadratmetern des tropischen Regenwaldes. Er kam auf 700 —
ebenso viele wie in ganz Nordamerika.
Der
Motor der Evolution 128
Dramatisch
ist die Zunahme der Artenzahl von Alaska bis zum tropischen Amerika auch bei
den Landvögeln: Sie steigt von 20 auf 600. George Stevens, der in beiden
Regionen lange gearbeitet hat, faßt den ersten Eindruck von diesen
Unterschieden so zusammen: »Wenn man durch Alaska fährt, ist man verblüfft
über die biologische Eintönigkeit. Ja, die geologischen Verhältnisse sind
faszinierend, aber Fauna und Flora sind immer gleich, wo man sich auch
befindet. In Costa Rica dagegen erkennt man schon bei den kleinsten
Geländeveränderungen große Unterschiede in den Lebensräumen.«4)
An
Land ist das Prinzip der zunehmenden Artenzahlen immer und immer wieder zu
erkennen. Über die Verhältnisse in den Meeren gab es bis vor kurzem nur
wenige Anhaltspunkte, denn dieser Lebensraum ist der Forschung natürlich viel
weniger zugänglich, insbesondere was das Leben in der Tiefsee angeht. Die
Tiefen der Ozeane, die früher als eine Art biologische Wüste galten,
beherbergen, wie wir heute wissen, ein gewaltiges Spektrum von Lebensformen.
Und seit man in den siebziger Jahren erstmals Lebensformen entdeckte, die ihre
Energie aus den durch vulkanisch-tektonische Aktivität entstandenen heißen
unterseeischen Quellen beziehen, weiß man auch, daß es dort höchst bizarre
Lebewesen gibt.
Das
neu entstandene Bild vom Leben in den Fluten ähnelt also verblüffend dem vom
Leben an Land; das bestätigte auch eine Gruppe von Wissenschaftlern aus den
Vereinigten Staaten, Schottland und Australien Ende 1993 in einer großen
Studie. Am größten ist die Artenvielfalt in der Nähe des Äquators, und
wenn man die Unterwassergebiete in höheren Breiten betrachtet, nimmt sie
immer weiter ab. »Der Rückgang der Artenvielfalt mit dem Breitengrad kam
unerwartet«, schrieben die Wissenschaftler, »denn man hatte angenommen, daß
die großflächige Verteilung der Umweltbedingungen an der Oberfläche die
Lebensgemeinschaften in größeren Tiefen nicht beeinflußt.«5
Die Tiefsee galt unter ökologischen Gesichtspunkten unabhängig von der
geographischen Breite immer als entsetzlich öde, und deshalb hatte man auch
angenommen, daß es keine großen Unterschiede in der Artenvielfalt
gibt.
Eine
endlose Zahl der schönsten Formen 129
Zu
Recht wiesen die Wissenschaftler daraufhin, daß der Artenschutz ihren
Befunden zufolge in den Meeren von ganz ähnlicher Bedeutung ist wie bei der
Zerstörung der Lebensräume an Land. So werden sich zum Beispiel die Suche
nach Rohstoffen, Ölförderung und Abfallbeseitigung in tropischen Gewässern
in ganz anderem Umfang auf die Artenvielfalt auswirken als in polnahen Meeren.
Mit
der Aussage, die Unterschiede der Artenzahl in der Tiefsee seien
»unerwartet« gewesen, sind wir bei der Frage nach den Ursachen dieses
weltweit gültigen Prinzips, und sie legt mindestens eine offenbar
vernünftige Annahme nahe: Die Abstufungen im Artenreichtum hängen an Land
unmittelbar mit Abstufungen wichtiger physikalischer Faktoren wie Temperatur
und Licht zusammen. Wir nehmen intuitiv an, daß die Lebensräume an Land und
in der Tiefsee sehr unterschiedlich aussehen müssen, und das stimmt auch. In
den Tiefen der Meere herrscht meist undurchdringliches Dunkel, und
Temperaturschwankungen werden abgemildert. An Land gilt das nicht.
Offenbar
ist irgendein grundlegender Mechanismus am Werk, der sich unter sehr
unterschiedlichen Umweltbedingungen auswirkt. Herauszufinden, was das für
ein Mechanismus ist, gestaltet sich nicht etwa deshalb schwierig, weil es
keine Hypothesen gäbe, sondern weil wir zu viele von ihnen haben. Man hat im
Laufe der Jahre zahlreiche Erklärungen angeboten, die einander häufig
unmittelbar widersprachen - ein heilsames Anzeichen, daß wir von einer
verläßlichen Antwort noch weit entfernt sind. Ich möchte einige dieser
Hypothesen genauer beschreiben.
Besonders
beliebt ist seit langem die sogenannte Zeittheorie. Danach herrschen in den
Tropen schon länger die gleichen Bedingungen wie in den gemäßigten
Klimazonen, und zwar wegen der immer wieder auftretenden Eiszeiten, die
gemäßigte Zonen wesentlich stärker beeinflussen. Deshalb, so die Vermutung,
stand in den Tropen mehr Zeit für die Ansammlung biologischer Vielfalt zur
Verfügung. Es gibt aber auf der Welt Gegenden, die von den Eiszeiten relativ
wenig betroffen waren; unter ihnen sind auch einige weiter nördlich gelegene
Geländeabschnitte, aber dort findet man nicht die höhere Artenvielfalt, die
man nach dieser Theorie erwarten würde.
Der
Motor der Evolution 130
Die
Zeittheorie ist offensichtlich nicht zu halten. Das gleiche gilt für die
»Produktivitätshypothese«, die ebenfalls lange Zeit im Schwange war.
Tatsächlich
sieht es so aus, als seien die Tropen von der Natur besonders gesegnet: Hier
herrschen angenehme Temperaturen, es gibt reichlich Licht und in vielen
Gegenden eine üppige Wasserversorgung. Aus unseren alltäglichen Erfahrungen
wissen wir, wie gut Pflanzen unter solchen Bedingungen gedeihen, und da Tiere
auf die Pflanzen angewiesen sind, ist eine große Vielfalt von Tieren und
Pflanzen möglich. Das stimmt, aber die Annahme, daß hohe Produktivität —
das heißt große Biomasse — auch notwendigerweise zu hoher Artenvielfalt
führt, stellt einen logischen Bruch dar. Warum sollten sich zahlreiche Arten
diese üppige Umwelt teilen und nicht nur wenige, die es sich in ihrem eigenen
Überfluß Wohlergehen lassen? (Die Nadelwälder des Nordens stellen eine
gewaltige Biomasse dar, bieten aber nur wenigen Arten Lebensraum; in
Graslandschaften dagegen ist die Biomasse unter Umständen gering, aber es
gibt zahlreiche Arten.) Die Schwäche dieser Theorie ist das Fehlen einer
erkennbaren Verbindung zwischen hoher Produktivität und der Entstehung vieler
Arten.
Interessant
ist dabei, daß die Zeithypothese den Tropen im Zusammenhang mit der Evolution
keine besonderen Eigenschaften zuschreibt, sondern nur mehr Zeit für die
Ansammlung von Arten postuliert. Die Produktivitätshypothese unterstellt
dagegen eine solche Besonderheit: Sie geht davon aus, daß die Tropen etwas an
sich haben, das häufiger zur Entstehung neuer Arten führt als in
gemäßigten Breiten. Wenn man durch Buschsavannen oder dichten, feuchten
Regenwald geht, fühlt man sich wie in einem Artenkaleidoskop, umgeben von
einer Fülle des Lebens auf allen Ebenen, wie in einem Fraktalmuster, das die
Kreativität der Evolution auszudrücken scheint. Die Annahme, daß die Tropen
den Strom des Lebens nähren und besonders häufig »eine endlose Zahl der
schönsten Formen« hervorbringen, scheint zu stimmen. Aber es gibt
auch eine andere Erklärung: Vielleicht sind die Tropen eine nachsichtigere
Umwelt, die Arten nicht so oft aussterben läßt wie die höheren Breiten.
Eine
endlose Zahl der schönsten Formen 131
Rein
gefühlsmäßig ist auch das eine reizvolle Annahme; in kühleren Gegenden
müssen Tiere und Pflanzen harte Winter überleben, und dabei können viele
von ihnen umkommen. Und Populationen, die lokal dezimiert werden, sind in
besonders harten Jahren eher vom Aussterben gefährdet. Der Kampf ums Dasein
scheint in den höheren Breiten mit ihren starken jahreszeitlichen
Schwankungen heftiger zu sein.
Bis
vor einiger Zeit konnte man nichts darüber sagen, ob die biologische Vielfalt
der Tropen durch besonders häufige entwicklungsgeschichtliche Neuerungen oder
durch eine geringere Aussterbehäufigkeit verursacht wird. Vor einigen Jahren
ging David Jablonski dieser ökologischen Frage jedoch nach, indem er auf die
Fossilfunde zurückgriff. Dabei ging er von folgender Überlegung aus: Wenn
sich an den Fossilien ablesen läßt, daß das erste Auftauchen neuer Arten in
den Tropen häufiger vorkommt als in gemäßigten Zonen, ist die Frage
beantwortet. Er untersuchte die Fossilien wirbelloser Meeresbewohner seit dem
Mesozoikum vor etwa 225 Millionen Jahren und erkannte dabei eindeutige
Anzeichen, daß Arten in den Tropen in größerer Zahl zum erstenmal
auftauchen. Im Juli 1993 schrieb er in Nature: »Dies ist ein
unmittelbarer Beleg, daß tropische Gebiete eine wichtige Quelle
entwicklungsgeschichtlicher Neuerungen waren und nicht nur ein
Rückzugsgebiet, in dem sich die Artenvielfalt wegen geringer
Aussterbehäufigkeit ansammelte.«6 Durch diesen wichtigen Befund war die
biologische Frage nun klarer umrissen: Was auch Besonderes an den Tropen sein
mag, es fordert tatsächlich die Evolution von Neuerungen.
Die
Biologen sind sich heute darüber einig, daß einer der wichtigsten Vorgänge,
durch den neue Arten entstehen, die sogenannte allopatrische Speziation ist.
Das heißt ganz einfach, daß Populationen einer vorhandenen Art aus
irgendeinem Grund voneinander getrennt werden, und in relativ kurzer Zeit
(einige tausend Jahre) sammeln sich dann so viele genetische Unterschiede und
Anpassungen an, daß daraus zwei selbständige Tochterarten werden. (Das war
die Kernaussage der im vorigen Kapitel beschriebenen
Puls-Fluktuationshypothese von Elisabeth Vrba.)
Der
Motor der Evolution 132
Wie
könnten die Umweltbedingungen der Tropen die allopatrische Speziation
fördern? Zu dieser Frage wurden zwei Vermutungen geäußert, die ich beide
beschreiben möchte.
Zuvor
möchte ich aber noch auf etwas anderes hinweisen: Es wäre naiv, sich die
Tropen als Gürtel einheitlich-üppiger Lebensräume vorzustellen, die jeweils
mit größerer Häufigkeit entwicklungsgeschichtliche Neuerungen ausspucken
als die gemäßigten Breiten. Wie die Biologen mittlerweile wissen, können
verschiedene Lebensräume die Populationen derselben Art in sehr
unterschiedlichem Ausmaß unterstützen, so daß diese Art in einer Gegend
vielleicht gut gedeiht, während sie woanders allmählich verschwindet.
Da
Tiere und (über das Ausstreuen der Samen) auch Pflanzen beweglich sind,
wandern aber Individuen aus dem ersten Lebensraum unter Umständen ständig in
den zweiten ein, so daß die Population in beiden recht einheitlich ist.
Derart unterschiedliche Lebensräume gibt es wahrscheinlich auch bei der
Artbildung; ein scheinbar einheitliches Stück Regenwald kann also in
Wirklichkeit ein Mosaik aus Lebensräumen sein, die unterschiedlich stark die
Bildung neuer Arten unterstützen.
Die
Ursache des Artbildungspotentials ist nach Ansicht vieler Biologen die
Stabilität der tropischen Lebensräume. »Wenn das Klima stabiler ist, kann
man auch mit stabileren Ressourcen rechnen«, erklärt Wallace Arthur, ein
Ökologe des Sunderland Polytechnic in England. »Deshalb können die Arten es
sich leisten, bei der Ernährung wählerischer zu sein — die Nahrungsnischen
sind kleiner, und sie überleben dennoch.«7
Wegen ihrer engen Nahrungsnischen bleiben solche spezialisierten Arten eher
auf kleine geographische Gebiete beschränkt, in denen die erforderlichen
Nahrungsquellen zur Verfügung stehen. Das bedeutet nicht nur, daß ein
bestimmtes Gebiet von viel mehr Arten besiedelt ist, sondern es begünstigt
auch die Neuentstehung von Arten. In dem gleichmäßigen tropischen Klima
können nach Arthurs Vermutung auch kleine Populationen überleben, und das
sogar am Rand ihres Verbreitungsgebietes; in den unwirtlichen nördlichen
Breiten dagegen werden solche Populationen eher verschwinden, und deshalb
entstehen dort auch seltener neue Arten.
Eine
endlose Zahl der schönsten Formen 133
Nach
dieser Vorstellung ist Stabilität die Triebfeder der Evolution, und
gleichzeitig bietet sie auch eine Umwelt, in der viele Arten nebeneinander
existieren können. Ist diese Vorstellung richtig?
Eine
in noch jüngerer Zeit entwickelte Hypothese sagt nein. Danach ist Instabilität
der Motor der Evolution. Während der siebziger und Anfang der
achtziger Jahre gab es unter den ökologisch orientierten Evolutionsforschern
eine heftige Debatte darüber, welche Kräfte die Struktur von
Lebensgemeinschaften beeinflussen. Zu Beginn der Diskussion herrschte die
Lehrmeinung, daß die Konkurrenz unter den Arten die Zusammensetzung der
Lebensgemeinschaften bestimmt, insbesondere wenn es darum geht, welche Arten
mit anderen zusammenleben können. Während der folgenden zehn Jahre wurde die
Konkurrenz als Faktor für die Struktur der Gemeinschaften zwar nicht völlig
verworfen, aber sie verlor stark an Bedeutung.
Großen
Einfluß auf diese Verschiebung der Ansichten hatte Joseph Connell, ein
Ökologe der University of California in Santa Cruz. Er berichtet, wie er als
überzeugter Anhänger des Konkurrenzgedankens nach Australien reiste, um die
Lebensgemeinschaften an Korallenriffen zu studieren, und wie ihn dort die
Natur eines Besseren belehrte: Ein Hurrikan wirbelte seine Untersuchungsstelle
durcheinander und fegte große Teile des Ökosystems hinweg. Für Connell war
es ein heilsamer Beweis, daß in der Natur neben der Konkurrenz auch andere
Kräfte wichtig sind. Zwar ist ein Hurrikan vielleicht eine besonders
dramatische und ungewöhnliche Umweltstörung, aber kleinere
Unregelmäßigkeiten sind häufiger. Und sie sind kreativer, wenn auch auf
indirekten Wegen.
In
Lebensgemeinschaften, die gegenüber Störungen unempfindlich sind,
übernehmen unter Umständen irgendwann wenige Arten die Vorherrschaft; bei
Belastungen jedoch, beispielsweise wenn Schneisen in ein Waldgebiet geschlagen
werden, haben andere Arten die Gelegenheit, zu einem Teil der Gemeinschaft zu
werden. Demnach, so kann man schließen, begünstigen wiederholte Belastungen
dieser Art die Evolution neuer Arten. Das dürfte der Intuition
widersprechen.
134
Eine
gutmütige, stabile Umwelt kann man sich eher als Wiege der Evolution
vorstellen, die neu entstehende Arten während ihres empfindlichsten Stadiums
erhält. In Wirklichkeit sieht es aber immer mehr danach aus, als seien
Belastung und Instabilität die Geburtshelfer neuer Arten.
Die
artenreichen Regenwälder des Amazonasbeckens waren zum Beispiel gewaltigen
Störungen ausgesetzt, insbesondere vor etwa 10.000 Jahren, als die letzte
Eiszeit zu Ende ging und die eher gleichförmige Jetztzeit begann. Zuvor, in
der kälteren Phase, gab es keinen Waldteppich, der den ganzen Kontinent
bedeckte, sondern in begünstigten Gebieten gediehen kleine Waldstücke, weil
dort das Mikroklima die an warmes Klima angepaßten Arten schützte. In der
Frage, wie eine solche Zerstückelung von Lebensräumen die Artbildung
begünstigt, bestehen schon seit langem Meinungsverschiedenheiten, die bis
heute nicht ausgeräumt sind. Nach einer Vorstellung waren die
Artenpopulationen in den einzelnen Waldstücken (auch Rückzugsgebiete
genannt) isoliert, so daß sie sich genetisch auseinanderentwickelten, ganz
ähnlich wie Populationen in anderen geographisch getrennten Regionen. Das
paßt gut zur Hypothese der allopatrischen Speziation, aber überzeugende
Hinweise, daß es solche Rückzugsgebiete gab, sind kaum zu finden. Vielleicht
waren schlicht Umweltstörungen der Motor der Evolution, eine kreative Umwelt
im Gleichgewicht zwischen völliger Stabilität und völliger Instabilität
— oder Chaos. (Im Amazonas-Regenwald trug die Maya-Hochkultur zur Störung
bei, weil ihre Angehörigen in manchen Gegenden große Waldflächen
abholzten.)
Die
Kreativität der Natur im Bereich zwischen Stabilität und Chaos erkennt man
auch in den Ozeanen. Hier herrschen seltsame Gesetzmäßigkeiten: Die größte
biologische Vielfalt der Meeresbewohner findet man in der Tiefsee. Dagegen
sind die Lebensgemeinschaften in der Nähe der Küsten relativ eintönig. Neue
Arten tauchen in beiden Bereichen auf, in größerer Zahl aber entstehen sie
in tieferen Gewässern. Die größten Neuerungen der Evolution — neue Arten,
die nicht nur Variationen vorhandener Themen darstellen, sondern ganz neue
Anpassungen besitzen — bilden sich jedoch häufiger in den küstennahen
Lebensgemeinschaften.
Eine
endlose Zahl der schönsten Formen 135
Es
scheint sich hier um einen realen Vorgang zu handeln, das heißt um die Folgen
größerer Neuerungen und nicht nur um das bevorzugte Überleben
entwicklungsgeschichtlicher Kuriositäten. Die Ursachen sind bisher jedoch
rätselhaft. Eines kann man allerdings feststellen: Die küstennahen
Gemeinschaften sind durch die Wellen stärkeren Störungen ausgesetzt als die
in der Tiefsee. Wieder sieht es so aus, als begünstigten Belastungen die
Entwicklung von neuem.
Wenn
Störungen die Geburtshelfer der Evolution sind, warum gibt es dann einen
Unterschied zwischen tropischen und gemäßigten Klimazonen? Einen möglichen
Grund nannte George Stevens vor kurzem: Tropische Arten sind häufig
Spezialisten, die sich an ein enges Spektrum von Umweltbedingungen angepaßt
haben; in gemäßigten Breiten müssen die Arten dagegen allgemeinere
Anpassungen besitzen, damit sie die jahreszeitlichen Temperatur- und
Lichtschwankungen überstehen können. Deshalb reagieren tropische Arten
empfindlicher auf Störungen, und das liefert der Evolution die Gelegenheit
zur Artbildung; die Arten der gemäßigten Regionen vertragen mehr und werden
deshalb weniger häufig in die entwicklungsgeschichtlich kreativeren
Randbereiche gedrängt.
Wie
steht es mit der Tiefsee, wo die Artenzahl sich mit den Breitengraden genauso
ändert wie an Land? Nach einer verbreiteten Vorstellung sind die Tiefen der
Meere ein Bereich des Endlos-Gleichen, in dem es kaum Veränderungen gibt und
der von den Wechselfällen der Umweltbedingungen abgeschnitten ist. Wenn das
stimmt, kann die Hypothese von den Störungen nicht zutreffen. Die Ökologen
John Gage und Robert May meinen jedoch in ihrem Kommentar zur Entdeckung der
abgestuften Artenzahl in der Tiefsee: »Vielleicht sind die Böden der Ozeane
weltweit doch nicht so einheitlich.«8
Möglicherweise stimmt das. Immerhin galt die Tiefsee noch vor nicht allzu
langer Zeit als biologische Wüste, und heute wissen wir, daß sie in
Wirklichkeit eine biologische Vielfalt hervorbringt, die an manchen Stellen
den tropischen Regenwäldern nahekommt.
Die
Verteilung der biologischen Vielfalt auf der Erde ist also höchst auffällig
und offenkundig. Und wie wir gesehen haben, schafft die Evolution »eine
endlose Zahl der schönsten Formen« auf Wegen, die sich auch heute noch dem
Verständnis der Biologen entziehen.
136
Die
zweitwichtigste Gesetzmäßigkeit in der biologischen Vielfalt auf der Erde
ergibt sich aus dem Vergleich der Lebensräume an Land und im Meer. Noch nicht
einmal 15 Prozent aller heute bekannten Arten leben in den Ozeanen, und von
diesen sind die meisten am Boden der Tiefsee oder kurz darüber zu Hause. Alle
anderen leben an Land. Besonders auffällig ist dieses Ungleichgewicht, weil
die Ozeane ja fast drei Viertel der Erdoberfläche bedecken. Offensichtlich
kann das Land also eine weitaus größere Vielfalt am Leben erhalten als das
Meer. Hier tut sich allerdings ein Widerspruch auf; er hat mit der Ebene in
der Hierarchie des Lebendigen zu tun, auf der man die beiden Bereiche
vergleicht.
Ich
habe bisher von den Arten gesprochen, also von der untersten Stufe der
biologischen Hierarchie. Vergleicht man die Lebensräume an Land und im Meer
auf der Ebene der Stämme, ergibt sich ein ganz anderes Bild als beim
Vergleich der Artenzahlen. In 32 der 33 Stämme des Tierreiches gibt es Arten,
die im Meer leben; an Land sind dagegen nur zwölf Stämme vertreten. Demnach
kommen 64 Prozent der Stämme ausschließlich im Meer vor, aber nur drei
Prozent sind ausschließlich an Land zu Hause. (Die übrigen haben Vertreter
an Land und in den Meeren.) An diesem Maßstab gemessen, bieten die Meere
einer weitaus größeren Vielfalt die notwendigen Lebensgrundlagen als die
trockenen Lebensräume. Mit anderen Worten: Im Meer findet man mehr Themen,
die jeweils nur wenige Variationen haben, an Land dagegen gibt es mehr
Variationen von weniger Themen.
Wie
für die mit dem Breitengrad abnehmende Artenzahl, so gibt es auch für die
auf Stamm- und Artenebene unterschiedliche Vielfalt an Land und im Meer
zahlreiche Erklärungsversuche — ein deutliches Anzeichen der Ungewißheit.
Selbst in einem sehr grundlegenden Sinn sind einfache Fragen manchmal schwer
zu beantworten, so zum Beispiel die, warum das Leben an Land so stark von den
Insekten beherrscht wird, während nur sehr wenige Arten dieser Gruppe im Meer
vorkommen.
Eine
endlose Zahl der schönsten Formen 137
Der
naheliegendste Grund, warum es in den Ozeanen mehr Stämme gibt, ist die
Tatsache, daß das vielzellige Leben dort in der kambrischen Explosion seinen
Anfang nahm. In dieser Phase oder kurz danach entwickelten sich alle heute
vorhandenen Stämme, und erst viel später wagten sich die ersten Lebewesen
auf das trockene Land. Alle heutigen Stämme hatten also die Gelegenheit, in
den Meeren Nachkommen zu hinterlassen; an Land bestanden ähnliche
Möglichkeiten nur für diejenigen, die sich an das Leben auf dem Trockenen
anpassen konnten. Das beantwortet aber nicht die Frage, warum es in den
Ozeanen weniger Abwandlungen der einzelnen Körperbaupläne gibt — immerhin
hatte das Leben dort gegenüber den landlebenden Formen einen Vorsprung von
100 Millionen Jahren. Es muß zwischen den beiden Lebensräumen einen
Unterschied geben, der den Motor der Evolution in einem davon schneller laufen
läßt.
Bestünde
die Tiefsee aus weiten, gleichförmigen Gebieten, wie man bis vor kurzem
angenommen hatte, wäre die geringere Artenvielfalt zu erklären. Eine in
Geländeform und Klima uneinheitliche Umwelt, die sich in relativ kurzen
Zeitabständen verändert, fordert die Evolution neuer Arten. Aber auch wenn
die Umwelt der Tiefsee sicher in mehrfacher Hinsicht ganz anders ist als die
an Land, dürfte sie doch viel weniger gleichförmig sein, als man früher
angenommen hatte.
In
den Tropen und bis vor nicht allzu langer Zeit auch in den gemäßigten Zonen
bieten die großen Wälder eine räumliche Komplexität, wie man sie im Meer
in der Regel nicht antrifft (außer an den Korallenriffen, die ebenfalls die
Heimat einer großen Artenvielfalt sind). Robert May, der sich eingehend mit
dieser Frage beschäftigt hat, räumt zwar ein, daß solche Paktoren eine
Rolle spielen könnten, aber er bleibt skeptisch: »Ich kann mir nur schwer
vorstellen, ... daß man damit das Verhältnis der Artenzahlen von 85 zu 15 an
Land und im Meer erklären kann.«9
Der
Motor der Evolution 138
Er
weist auch auf einen seltsamen Unterschied zwischen tropischen Wäldern und
Korallenriffen hin, von denen man oft sagt, sie entsprächen einander in ihren
jeweiligen Bereichen. »Das Korallenriff strotzt von auffälligen Tieren ...
im tropischen Regenwald dagegen bekommt man die Wirbeltiere kaum einmal zu
Gesicht, und selbst die Wirbellosen sind nicht so leicht zu erkennen.«10
Den Grund für diesen Unterschied kennt niemand.
Man
hat noch mit vielen anderen Hypothesen versucht, den unterschiedlichen
Artenreichtum an Land und im Meer zu erklären; die vielleicht
vielversprechendste hat mit der Größe und dem Verbreitungsgebiet der Arten
zu tun. Meeresbewohner sind im Durchschnitt kleiner als landlebende Arten,
aber ihr Verbreitungsgebiet ist größer. Und eine größere geographische
Verbreitung bedeutet — jedenfalls in der Regel — eine insgesamt geringere
Artenzahl. Vor dem Hintergrund der Evolutionsökologie hört sich diese
Erklärung plausibel an, aber sie ist zugegebenermaßen Spekulation.
Von
der weltweiten Verteilung der Artenvielfalt möchte ich jetzt zu ihrer
Zusammensetzung zurückkehren und die Frage stellen: Wie viele Arten gibt es
heute auf der Erde?
Auf
diese einfache Frage hat Robert May eine einfache Antwort: »Wir sind uns in
der Größenordnung einer Zehnerpotenz unsicher, mit wie vielen Arten wir den
Globus teilen.«11
Die meisten Schätzungen bewegen sich zwischen fünf und 50 Millionen, ja
manche gehen sogar bis 100 Millionen. Der Grund für diese gewaltige
Unsicherheit? Kaum ein Biologe hat jemals versucht, eine Antwort zu finden,
und wer es wagte, war entmutigt von den Schwierigkeiten, die sich dabei
ergeben.
Es
ist schon bemerkenswert, daß wir in unserer modernen Welt und mit unserer
Besessenheit, alles zu messen, nur so ungenaue Vorstellungen über den Stoff
der Natur haben, mit dem wir so eng verflochten sind und von dem wir letztlich
abhängen. Für die Zahl der Sterne in unserer Galaxis, der Milchstraße,
haben wir eine gute Schätzung: Es sind ein paar hundert Milliarden. Wir
wissen, wie viele Nukleotide den genetischen Bauplan des Menschen bilden: drei
Milliarden.
Eine
endlose Zahl der schönsten Formen 139
Und
wir können auf wenige Stunden genau berechnen, wann ein Komet auf den Jupiter
stürzen wird, wie es am 16. Juli 1994 um 22 Uhr Mitteleuropäischer
Sommerzeit geschah. Und doch können wir die heutige Artenvielfalt nicht mit
einer gesicherten Zahl benennen. Es liegt nicht daran, daß man nicht wüßte,
wie man sie ermittelt, sondern es fehlt schlicht der Wille. Die Regierungen
haben viele hundert Millionen Dollar in die systematische Untersuchung der
Sterne gesteckt, aber nur ein winziger Bruchteil dieser Summe floß in die
systematische Untersuchung der Natur hier bei uns auf der Erde.
Die
Suche nach Ordnung in der Natur begann eigentlich mit Aristoteles, aber bis
die Naturgeschichte zu einer angesehenen Disziplin der abendländischen
Wissenschaft wurde, sollten noch fast zwei Jahrtausende vergehen. Zum
wichtigsten Anliegen der neuen Wissenschaft wurde die Einteilung der Pflanzen
und Tiere, anfangs im Zusammenhang mit der Absicht, die Ergebnisse von Gottes
Schöpferkraft vorzuführen. Das heutige Klassifikationssystem begründete
Carl von Linnc im 18. Jahrhundert mit seinem Systema naturae. Er
listete etwa 9000 Tier- und Pflanzenarten auf und bediente sich dabei eines
Systems, in dem ihre Verwandtschaftsbeziehungen deutlich wurden (natürlich im
Sinne der Schöpfung und nicht der geschichtlichen Entwicklung). Dieser
bahnbrechende, aber noch bescheidene Versuch einer systematischen Erforschung
der Natur kam erst ein volles Jahrhundert, nachdem Isaac Newton die
Gravitationsgesetze formuliert hatte, auf die sich die Berechnungen der
jüngsten Kollision zwischen Komet und Jupiter gründeten.
Seit
Linnes Zeit ist die Zahl der beschriebenen Arten natürlich beträchtlich
angewachsen; derzeit liegt sie bei ungefähr 1,4 Millionen. Ich sage
»ungefähr«, weil es keine zentrale Sammelstelle für alle Beschreibungen
von Arten gibt, so daß die Zahl immer nur eine Schätzung sein kann. Das ist
besonders verwunderlich: Für die DNA-Sequenzen, die in den Labors auf der
ganzen Welt analysiert werden, gibt es eine zentrale Sammelstelle, aber nicht
für die Lebewesen, aus denen man das genetische Material gewinnt.
Der
Motor der Evolution 140
Etwa
85 Prozent der bekannten Arten leben an Land, und die Mehrzahl von ihnen,
insgesamt ungefähr 850.000, sind Gliederfüßer (Insekten, Spinnen und
Krebstiere). Die meisten Gliederfüßerarten sind Insekten, und von diesen
sind etwa die Hälfte Käfer, eine Tatsache, die den britischen Biologen J.B.S.
Haidane zu einem berühmten Ausspruch angeregt haben soll. Als ihn einige
geistliche Herren einmal fragten, was er bei seiner Naturforschung über Gott
erfahren habe, soll Haidane geantwortet haben, es deute alles darauf hin, daß
Er »eine außerordentliche Vorliebe für Käfer« habe. Die Geschichte ist
zwar nicht belegt, aber sehr treffend, und wenn Studien aus jüngerer Zeit
recht haben, hat man die Wirklichkeit bisher sogar noch unterschätzt.
Bei
den 300.000 bekannten Pflanzenarten handelt es sich in der Mehrzahl um
Blütenpflanzen. (Daß gerade Insekten und Blütenpflanzen so artenreich sind,
ist sicher kein Zufall, denn beide Gruppen haben sich in den letzten paar
hundert Millionen Jahren gemeinsam entwickelt.) Außerdem kennt man etwa
69.000 Pilzarten und ungefähr ebenso viele Arten einzelliger Lebewesen. Zu
der letztgenannten Gruppe gehören nur ungefähr 5000 beschriebene
Bakterienarten. Die Kategorie der Wirbeltiere, die unsere Aufmerksamkeit am
stärksten erregen, vereinigt etwa 40.000 Arten auf sich, darunter 4000
Säugetiere und 9000 Vögel; der Rest verteilt sich auf Reptilien, Amphibien
und Fische.
Wer
nun aber meint, die 4000 Säugetierarten stünden gegenüber der Zahl der
verschiedenen Bakterien doch gar nicht so schlecht da, sollte seinen Stolz
einen Augenblick lang zügeln. Vor kurzem analysierte eine norwegische
Wissenschaftlergruppe die Bakterienwelt in einem Gramm Erde aus einem
Buchenwald und in einer ähnlichen Sedimentmenge von der norwegischen Küste.
In beiden Fällen fanden die Wissenschaftler ungefähr 5000 Arten, und zwar
ohne Überschneidungen zwischen den beiden Bodenproben. Die bisher
beschriebenen 5000 Bakterienspezies sind also offenbar nur ein winziger
Bruchteil der wirklichen Zahl.
Aber
die Liste der bisher bekannten Arten führt nicht nur zu einer gewaltigen
Unterschätzung der tatsächlichen Artenzahl, sondern sie ist auch in
mehrfacher Hinsicht unausgewogen. Erstens spiegelt sich in ihr das durchaus
nicht unnatürliche Interesse der Menschen an behaarten und gefiederten
Geschöpfen wider.
141
|
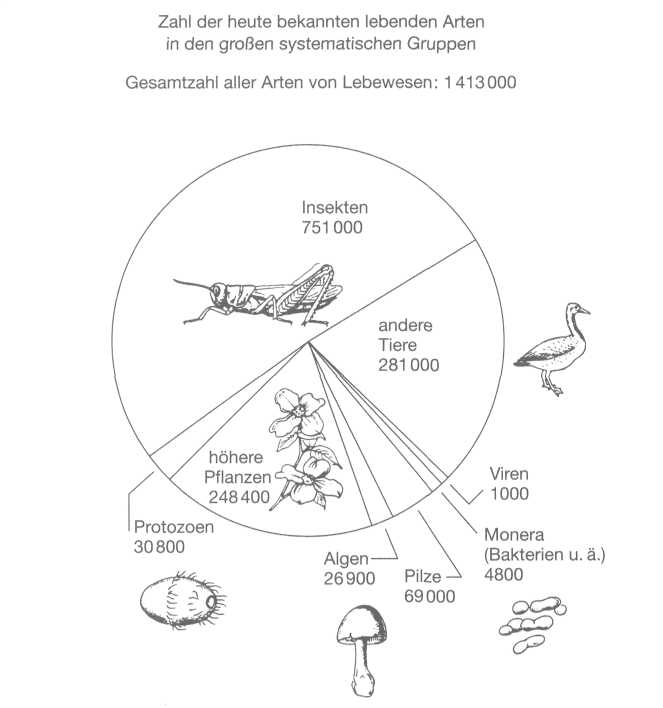 |
Die
beherrschenden Gruppen in der Vielfalt der heute bekannten Lebensformen
sind Insekten und Pflanzen, aber bei den Bakterien, Pilzen und anderen
kaum untersuchten Gruppen ist eine Riesenzahl von Arten noch nicht
entdeckt.
Die
Gesamtzahl aller Arten liegt irgendwo zwischen zehn und 100 Millionen.
(Nachgedruckt
mit freundlicher Genehmigung der Verleger von The Diversity of Life von
Edward O. Wilson; Cambridge, Mass.: The Belknap Press of Harvard
University Press, © 1992 by Edward O. Wilson.)
|
Der
Motor der Evolution 142
Mit
Vögeln und Säugetieren beschäftigen sich weitaus mehr Systematiker als
beispielsweise mit Insekten, Fadenwürmern oder Bakterien. Deshalb werden zwar
immer noch jedes Jahr neue Vogel- und Säugetierarten entdeckt, aber es sind
nur wenige, und die Gesamtzahl dürfte letztlich nicht wesentlich über der
heute bekannten liegen. Für die übrige Natur — das heißt für ihren
größten Teil — gilt das nicht, wie man an dem Beispiel mit den Bakterien
deutlich erkennt. Eine zweite Verzerrung ergibt sich, weil man sich vorwiegend
auf die gemäßigten Klimazonen des Nordens konzentrierte, in denen die
meisten Systematiker (Spezialisten für die Einteilung der Lebewesen)
arbeiten. Die überwiegende Mehrheit aller Arten lebt in den Tropen, und
dennoch kommen auf jede Art, die man dort kennt, zwei andere aus höheren
nördlichen Breiten.
Einen
der ersten Versuche, eine wissenschaftlich begründete Schätzung für die
Zahl der Insektenarten abzugeben, unternahm der britische Ökologe Carrington
Williams. Er veröffentlichte 1964 ein Buch mit dem Titel Patterns
in the Balance of Nature (»Muster im Gleichgewicht der Natur«). Darin
gelangte er mit einer Kombination aus lokalen Beobachtungen und Hochrechnungen
zu einer Zahl von drei Millionen. In den darauffolgenden 20 Jahren sammelten
die Freilandbiologen — meist in unabhängigen Untersuchungen — weitere
Erkenntnisse über die unterschiedlichsten Lebensräume, darunter auch zuvor
völlig unbekannte wie den Boden der Tiefsee. Daraufhin stiegen die
Schätzungen für die Gesamtzahl der Arten auf mindestens zehn Millionen.
Dann
folgte eines der dramatischsten Ereignisse in der biologischen Systematik:
Terry Erwin von der Smithsonian Institution gab 1982 bekannt, es gebe
wahrscheinlich allein 30 Millionen Insektenarten, die meisten davon im
Kronendach der tropischen Regenwälder. Zu dieser Schätzung gelangte er,
nachdem er die Insektenpopulation an einer großen Baumgruppe im Regenwald
Panamas untersucht hatte. Er hatte zu diesem Zweck Insektengifte in den
Baumkronen versprüht und dann die toten Tiere gezählt, die auf den Boden
fielen.
143
|
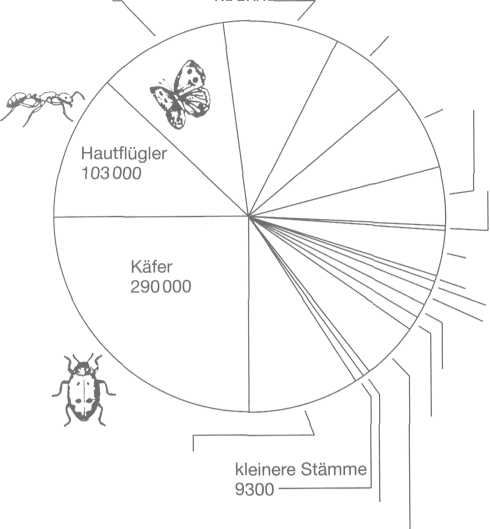 |
Bei
den Tieren, die der Wissenschaft bekannt sind, handelt es sich in der
überwältigenden Mehrheit um Insekten. Wegen dieses Übergewichts sind
die meisten Tierarten Landbewohner; die meisten Stämme dagegen
(Stachelhäuter usw.), welche die höchsten Klassifikationseinheiten
darstellen, leben im Meer.
(Nachgedruckt
mit freundlicher Genehmigung der Verleger von The Diversity of Life von
Edward O. Wilson; Cambridge, Mass.: The Belknap Press of Harvard
University Press, © 1992 by Edward O. Wilson.)
|
Der
Motor der Evolution 144
Es
war ein kühnes Unternehmen, aber obwohl es wissenschaftlich von großer
Bedeutung war, blieb es weitgehend unbemerkt. Edward Wilson meinte dazu einige
Jahre später: »Wenn die Astronomen jenseits des Pluto einen neuen Planeten
entdecken würden, stünde die Nachricht auf der ganzen Welt in den
Schlagzeilen. Bei der Entdeckung, daß die Welt des Lebendigen reichhaltiger
ist als vermutet, ist das nicht der Fall, obwohl sie für die Menschheit viel
wichtiger ist.«12
Seit
Erwin seine Entdeckung bekanntgab, wächst das Interesse an der nicht
beantwortbaren Frage. Auch diesmal war es kein koordiniertes Unternehmen,
sondern es kam von Biologen, die unabhängig voneinander auf ihren eigenen
Forschungsgebieten arbeiteten. So behauptet beispielsweise David Hawksworth
vom International Mycologieal Institute im englischen Kew, die derzeitige
Schätzung von 69.000 Pilzarten liege um mindestens das Zwanzigfache zu
niedrig. Wie er zeigen konnte, kommen in den Lebensräumen Europas auf jede
Art von Gefäßpflanzen etwa sechs Pilzarten; und da man ungefähr 300.000
solche Pflanzenspezies kennt, gibt es demnach vermutlich 1,8 Millionen
Pilzarten. Und wenn noch mehr Pflanzenarten entdeckt werden, dürfte auch
diese ohnehin hohe Zahl weiter steigen. Ähnliches gilt für die Nematoden, so
Peter Hammond vom Museum of Natural History in London. Heute kennt man etwa
15.000 Arten der winzigen Fadenwürmer. Die Gesamtzahl der Arten dieser
allgegenwärtigen Parasiten, die auf Tieren und Pflanzen sowie selbständig in
Salz- und Süßwasser leben, dürfte sich nach Hammonds Berechnungen auf
ungefähr 300.000 belaufen.
Solchen
und anderen Schätzungen zufolge liegt die Gesamtzahl der Arten bei fast 50
Millionen, und selbst diese Zahl ist vielleicht noch zu niedrig. Robert
May kommt in einer Berechnung, die er »kühn, aber nicht unbegründet«
nennt, sogar auf 100 Millionen Arten. Er stützt sich dabei auf die
Vorstellung, daß »auf jede Spezies von den beiden artenreichsten
Organismengruppen, den Gliedertieren und den Gefäßpflanzen, zumindest ein
Fadenwurm, ein Protozoon, ein Bakterium und ein Virus kommen, die allein bei
ihr schmarotzen«.13)
145

Ein
Nest des Bleßhuhns Fulica chloropus (Linnaeus). Aus dem Museum Rouen.
Eine
endlose Zahl der schönsten Formen 146

Ameisen
kehren vom Kampf heim.
147

Die
Vogelspinne Mygale avicularia tötet einen Kolobri. (Maria Sibylla Merian)
148

Der
Laufkäfer Calosoma Inquisitor verfolgt einen Bombardierkäfer (Brachinus
crepi-tans), der sich beim Rückzug verteidigt.
In
jedem Fall dürfte heute klar sein, daß die meisten Arten, seien es nun
insgesamt 30, 50 oder 100 Millionen, in den Tropen
leben und sehr unauffällig sind. Die Welt der großen Wirbeltiere und
Pflanzen, die wir täglich erleben, stellt nur einen Bruchteil der Vielfalt
des Lebendigen dar. Wir erkennen die Gestalt der Vielfalt mit relativ
wenigen großen und vielen kleinen Organismen, und wir wissen, daß dies unter
anderem mit dem Energiefluß in den ökologischen Gemeinschaften zu tun hat.
Aber das Ausmaß der Vielfalt können wir aus solchen Grundregeln nicht
ableiten; es gibt in Ökologie und Evolutionsbiologie keine theoretische
Grundlage für die Behauptung, die Erde könne bei der derzeitigen Verteilung
der Kontinente eine, zehn, 30, 50 oder 100 Millionen Arten am Leben erhalten.
Alle
diese Angaben über die Artenzahl sind mit großen Unsicherheiten behaftet,
denn sie gründen sich immer auf eine Art Hochrechnung aufgrund von
Freilandmessungen. Manche Berechnungen werden sich zwangsläufig als falsch
erweisen, wenn sich herausstellt, daß die Verhältnisse zwischen den
Artenzahlen (beispielsweise zwischen Pilzen und Pflanzen) in verschiedenen
Gegenden der Erde unterschiedlich sind.
149
Dennoch
ist es der vernünftigste Weg, um zu Angaben über Artenzahlen zu gelangen. Die
Biologen haben ungefähr 230 Jahre gebraucht, um drei Viertel von einer
Million Insektenarten zu identifizieren und zu beschreiben; wenn es
tatsächlich, wie Erwin schätzt, 30 Millionen gibt, hätten die
Insektenspezialisten bei Beibehaltung der bisherigen Arbeitsweise noch
Beschäftigung für etwa 10.000 Jahre. Die vollständige Erfassung aller
Pflanzen Nord- und Südamerikas würde die Systematiker — ebenfalls bei der
bisherigen Arbeitsgeschwindigkeit — etwa 400 Jahre lang mit Beschlag
belegen, so eine Schätzung des Leiters der botanischen Gärten in Kew,
Ghilean Prance.
Die
Insekten- und Pflanzenspezialisten zählten nicht einfach nur Arten, sondern
sie beschrieben sie auch. Hinter jedem Katalogeintrag verbirgt sich eine
einzigartige Lebensform, ein Erbe aus Hunderten von Jahrmillionen der
Evolution, in der wir nur ein kleiner Teil sind. Aber die Zahl der Einträge
ist jämmerlich klein, und sie zu steigern, ist eine gewaltige Ausgabe,
gemessen an den Mitteln, welche die abendländische Wissenschaft ihr bisher
gewidmet hat. Es wirft schon ein betrübliches Licht auf den Wert, den wir der
gewaltigen Vielfalt hier auf Erden beimessen, wenn May sagen muß: »Wir sind
uns in der Größenordnung einer Zehnerpotenz unsicher, mit wie vielen Arten
wir den Globus teilen.«
Sollen
wir, wie Edward Wilson fordert, »nichts Geringeres anstreben als eine
vollständige Zählung, einen kompletten Katalog des Lebens auf der Erde«?14)
Ein solches Unternehmen wäre kostspielig, sicher, aber der Aufwand wäre
geringer als der für die Sequenzierung des menschlichen Genoms, die 150
Millionen Dollar im Jahr verschlingt, oder für den Bau einer Weltraumstation,
die insgesamt auf etwa 30 Milliarden Dollar kommt.
Als
Direktor der kenianischen Naturschutzbehörde, die ein knappes Jahr nach
meiner Amtsübernahme in »Naturschutzdienst« umgetauft wurde, erlebte ich
Tag für Tag, was es kostet, auch nur die auffälligsten Arten zu schützen.
Und ich kann deutlich erkennen, daß eine Summe von beispielsweise 100
Millionen Dollar vielen Ländern helfen würde, die sich angesichts einer
wachsenden Bevölkerung mit dem Schutz der natürlichen Tierwelt
herumschlagen.
Aber
Wilsons ehrgeiziges Ziel ist
lohnend: lohnend für die Wissenschaft und lohnend für die Menschheit. Als
krönender Abschluß der Evolution, als vernunftbegabte Spezies haben
wir die moralische Verpflichtung, soviel wie möglich über die »endlose
Zahl der schönsten Formen« zu wissen, mit denen wir unsere Erde teilen.
Der
Goliathkäfer Golianthus giganteus
150-151
#
www.detopia.de
^^^^
Richard Leakey 1995