Start Weiter
Teil 3 Gleichgewicht
der Natur ? Leakey 1995
Ökologische Lebensgemeinschaften sind nichts
Freundlich-Harmonisches. Sie werden von vielerlei Kräften geformt, manche
davon dem Chaos, andere dem Zufall entsprungen. Vor allem sind sie in
ständigem, dynamischem Wandel begriffen. Die
Menschen hatten in der historisch belegten Vergangenheit großen Einfluß auf
solche Lebensgemeinschaften; daran wird deutlich, wie leicht Ökosysteme
durch neu hinzukommende Arten gestört werden. Die
heutige Misere der Elefanten zeigt nicht nur, welche Auswirkungen wir auf
die Natur haben können, sondern sie offenbart auch, welche Herausforderung
es bedeutet, komplexe ökologische Systeme zu schützen.
9 Stabilität und Chaos in der Ökologie
183-208
Auf
einer Reise quer über den Globus vom Pol zum Äquator würde man das
erkennen, was zusammenfassend als »die unendliche Vielfalt der Natur«
bezeichnet wurde. Von der eisigen Tundra zu den Almwiesen der Gebirge, von
Wäldern und Feldern der gemäßigten Breiten zu den tropischen Regenwäldern
und Savannen würde man ein riesiges Spektrum der verschiedensten
ökologischen Gemeinschaften erleben. Diese Unterschiede sind für
Nordamerikaner und Europäer ein wichtiger Beweggrund, jedes Jahr in großer
Zahl zu uns nach Kenia zu kommen.
Der
Kontrast zwischen den Lebensgemeinschaften von Pflanzen, Insekten, Vögeln und
anderen Tieren in der Heimat und in den bereisten Gebieten ist gewaltig. Das
liegt nicht nur daran, daß die Zahl der Arten von den hohen zu den niederen
Breiten zunimmt; auch das Wesen der Arten in den Lebensgemeinschaften ändert
sich. (In tropischen Ökosystemen findet man keine Eisbären, und große
Primaten — vom Menschen einmal abgesehen — gelten nicht als natürlicher
Bestandteil der Tierwelt in gemäßigten und polaren Gebieten.)
Diese
großräumige Verteilung ergibt sich natürlich zum Teil aus der Anpassung der
Arten an die örtlichen Umweltbedingungen, insbesondere an Temperatur und
Luftfeuchtigkeit. Eine lokale ökologische Lebensgemeinschaft ist also
eine Ansammlung von Arten, denen bei allen individuellen Unterschieden eine
Anpassung gemeinsam ist: die Anpassung an die Gegebenheiten der örtlichen
Umwelt.
Um
eine — zwar nicht unendliche, aber doch großartige — Vielfalt zu erleben,
muß man aber kein Globetrotter sein. Wo man sich auch auf der Erde befindet,
immer ist man von ökologischen Gemeinschaften umgeben, die sich voneinander
unterscheiden, und zwar manchmal sehr grundlegend.
Ich
habe bereits die dramatischen Gegensätze zwischen den Ökosystemen des
Rift-Tals in Kenia beschrieben. Auch dort erkennt man ohne weiteres die
Ursachen der Vielfalt: Von der Sohle des Tals bis zu den höchsten Punkten
seiner Flanken herrschen unzählige Mikroklimata, die den verschiedensten
Arten höchst gegensätzliche Lebensbedingungen bieten. Evolution und
Anpassung wirken in allen Größenordnungen und schaffen Verteilungsmuster
aller Größenordnungen.
Die
Ökologie hat unter anderem das Ziel, »die Muster der natürlichen
Ökosysteme aufzuspüren und die Kausalprozesse zu erklären, die ihnen
zugrunde liegen«, so eine Formulierung des Ökologen John Wiens von der
University of New Mexico. Der wichtigste Prozeß bei dem, was ich bisher
beschrieben habe, ist die Anpassung an örtliche Gegebenheiten — das
jedenfalls habe ich nahegelegt. Aber wer schon einmal aufmerksam durch einen
Wald oder über eine Wiese gestreift ist, der weiß, daß Natur nichts
Gleichförmiges, sondern ein Flickenteppich ist.
Eine
Zeitlang sieht man auf dem Spaziergang immer wieder eine Baumart, die später
völlig fehlt; eine Blumenart kommt am südlichen Ende der Wiese nur selten
vor, im Norden ist sie häufig. Dieser Flickenteppich der Natur ist ein
Flickenteppich ähnlicher, aber doch unterschiedlicher ökologischer
Gemeinschaften. Welche Vorgänge haben diese Verteilung geprägt? In
den Umweltbedingungen an den beiden Enden der Wiese gibt es keine erkennbaren
Unterschiede, warum also sind die Ökosysteme nicht gleich?
Vielleicht
übersieht auch der aufmerksamste Beobachter, daß es doch entscheidende
Unterschiede gibt, beispielsweise in der chemischen Zusammensetzung des Bodens
oder im Grundwasserspiegel. Solche Erklärungen sind bei manchen Ökologen
sehr beliebt; so schrieb Seth Rice, Biologe an der University of North
Carolina, in einem wichtigen Übersichtsartikel: »Alle Umweltbedingungen in
allen Ökosystemen sind räumlich und zeitlich wandelbar. Das mosaikartige
Bild der Umwelt ist die Folge allgegenwärtiger physikalischer und chemischer
Abstufungen.«1
Mit anderen Worten: Die biologischen Verteilungsmuster werden auf dem Wege der
lokalen Anpassung von den Verteilungsmustern der physikalischen Umwelt
bestimmt und spiegeln diese wider.
184/185
Stimmt
das wirklich? Intuitiv erscheint es plausibel und sogar offenkundig, daß
Ökosysteme auf diese Weise geformt werden, aber in den letzten Jahren hat
sich gezeigt, daß hier auch andere Kräfte mitwirken, Kräfte, die weniger
leicht zu erkennen sind und auf den ersten Blick sogar unplausibel erscheinen.
Das
Gebiet, auf dem wir uns hier bewegen — die Ökologie der
Lebensgemeinschaften —, stellt vermutlich einige der wichtigsten und am
wenigsten lösbaren wissenschaftlichen Probleme. Im Kern geht es um eine
einzige einfache Frage: Wie wird eine ökologische Gemeinschaft zu dem, was
sie ist? Eine Antwort, die sich lange Zeit großer Beliebtheit erfreute,
lautet: Die Gemeinschaft ist so, weil sie so sein muß, weil die örtlichen
Gegebenheiten es so vorschreiben. Und, was noch wichtiger ist: Die Mitglieder
der Gemeinschaft sind dieser Annahme zufolge gemeinsam so eng an die
Gegebenheiten angepaßt — und damit auch so abhängig voneinander —, daß
eine Gemeinschaft mit anderer Artenzusammensetzung nicht überleben könnte.
In dieser Form ist die Aussage vielleicht ein wenig streng formuliert, aber
sie gibt das Wesentliche eines großen Teils der derzeitigen ökologischen
Denkweise wieder. Im Zusammenhang mit dieser Ansicht über den grundlegenden
Aufbau ökologischer Gemeinschaften steht auch die Vorstellung vom
»Gleichgewicht der Natur«.
Auch
dieser Begriff und seine Folgerungen erscheinen intuitiv vernünftig und sogar
beruhigend. Wenn die Ökosysteme so sind, wie sie sein müssen, dann folgt
daraus, daß die Natur sie schnell wiederherstellt, wenn sie aus irgendeinem
Grund gestört werden.
Vor
einigen Jahren faßte
Fairfield Osborn,
der Sohn
von Henry Fairfield Osborn, diese Einstellung in seinem Buch Our
Plundered Planet so zusammen: »Natur mag etwas Schönes sein und ist
sogar eine Symphonie, aber über und unter und in ihrem unwandelbaren Wesen,
ihren Abständen, ihrer scheinbaren Ruhe und Unveränderlichkeit arbeitet eine
aktive, zielgerichtete, koordinierte Maschine.«
Die
Maschine hält die Gemeinschaften in ihrem Gleichgewichtszustand. Der Ausdruck
»Gleichgewicht der Natur« wurde in Ökologen- und Laienkreisen zu einer
eindringlichen Metapher für die
beobachtete grundlegende natürliche Harmonie, die das Gefühl erweckt, die
Welt, die wir erleben, sei richtig.
*detopia:
Osborn bei detopia
186
Die
Folge war, daß der Ökologie eine Zeitlang der Geruch des Mystischen
anhaftete. Aber selbst als die Mystik vor ein paar Jahrzehnten verschwand,
blieb der Ausdruck erhalten. Nachdem man ihn aller Vorstellungen von Absicht
entkleidet hatte, bezeichnete er die Fähigkeit einer ökologischen
Gemeinschaft, sich einer Störung zu widersetzen oder sich von ihr zu erholen,
eine Eigenschaft, die man nun auch in den objektiveren Begriff Stabilität faßte.
Aber ob man nun »natürliches Gleichgewicht« oder »Stabilität« sagt: Wie
Stuart Pimm anmerkt, »haben beide Begriffe unverkennbar etwas Verwaschenes«.2
Wie
sich eine ökologische Gemeinschaft zusammenfindet, ist ebenso eine immer noch
unbeantwortete Frage wie die nach dem Verhalten und den Eigenschaften der
bestehenden Gemeinschaften. Das sind unglaublich komplizierte Themen, denn sie
enthalten viele Variablen (das heißt einzelne Arten), die auf vielerlei Weise
in Wechselwirkung treten können — und das Ganze spielt sich unter häufig
recht turbulenten physikalischen Umweltbedingungen ab. Deshalb gibt es ein
riesiges, eigentlich sogar unendliches Spektrum möglicher Verteilungsmuster,
und herauszufinden, warum manche davon sich ausbilden und andere nicht, ist
eine entmutigende Aufgabe.
Das
alles mag sehr theoretisch klingen, und einerseits besteht auch der starke
Drang zu verstehen, wie die Natur funktioniert und woher die biologische
Vielfalt stammt, zu der auch wir gehören. Aber diese Kenntnisse sind auch
unentbehrlich für unsere Bestrebungen, die biologische Vielfalt zu schützen
und den unendlichen Reichtum der Natur zu erhalten. So sollten wir zum
Beispiel nicht nur in der Lage sein, die Artenzusammensetzung der
Lebensgemeinschaften zu erklären, sondern wir müssen auch verstehen, warum
die Populationsgröße der einzelnen Arten innerhalb der Gemeinschaften
schwankt. Wir müssen in Erfahrung bringen, warum manche Gemeinschaften für
Störungen — besonders für solche, die vom Menschen verursacht werden —
anfällig sind, während andere ihnen widerstehen.
187
Wir
müssen wissen, warum manche Gemeinschaften sich schnell von Zerstörungen
erholen, während das bei anderen sehr lange dauert. Wir müssen herausfinden,
warum fremde Arten in manche Gemeinschaften leicht einwandern können, in
andere dagegen nicht, und wir müssen die Folgen solcher Verschiebungen
einschätzen. Wir müssen verstehen, welche Arten vom Aussterben bedroht sind
und welche in ihren jeweiligen Lebensgemeinschaften eine so große Bedeutung
haben, daß ihr Verschwinden zu einer Lawine weiterer Aussterbeereignisse
führt. Alle diese Fragen nehmen in den Lehrbüchern der Ökologen und den
Handbüchern der Naturschützer einen gleichermaßen wichtigen Platz ein.
Am
Anfang des vorliegenden Kapitels steht eine genauere Untersuchung einer in
jüngster Zeit gewonnenen Erkenntnis, die der Intuition zu widersprechen
scheint. Sie betrifft die Gründe, warum die Populationen der Arten in der
Weise schwanken, wie man es beobachtet: manchmal regelmäßig, manchmal heftig
und unberechenbar. Das führt uns unmittelbar zu der Einsicht, daß die Natur
meist keineswegs im Gleichgewicht ist, sondern sich chaotisch verhält. Der
eine oder andere mag ein derart beunruhigendes Bild von der Natur vielleicht
nicht akzeptieren, denn es scheint aller grundlegenden Harmonie zu entbehren.
Anschließend
werde ich erörtern, wie man in der Ökologie einige Vorgänge, durch die sich
Lebensgemeinschaften bilden, kennengelernt hat; zu diesem Zweck waren die
Wissenschaftler meist gezwungen, umfangreiche Computermodelle zu entwickeln.
Wir werden sehen, daß ökologische Gemeinschaften einen eigenen Geist zu
haben scheinen, denn sie verbessern sich im Laufe der Zeit und werden
gegenüber dem Eindringen fremder Arten immer widerstandsfähiger. Ich werde
auf die Dynamik dieses Eindringens zu sprechen kommen — auf die Frage, was
einen erfolgreichen Eindringling ausmacht und wovon die Auswirkungen solcher
Ereignisse abhängen. Das ist ein wichtiges Thema für den Naturschutz. Und
schließlich werde ich einen warnenden Vorschlag machen, wie man als
Naturschützer für die Stabilität der Ökosysteme sorgen sollte — nämlich
indem man ihnen gestattet, sich zu wandeln.
188
Auch
das widerspricht wie so vieles, das man über die Ökologie der
Lebensgemeinschaften erfahren hat, der Intuition. Die Natur ist nicht immer
so, wie sie zu sein scheint.
Im
National Museum of Natural History der Smithsonian Institution in Washington
gibt es eine Vitrine, die jedem Besucher einen kalten Schauer über den
Rücken laufen läßt: Sie zeigt eine verschmutzte Küche, die über und über
mit Küchenschaben bedeckt ist — es sind Hunderttausende, vielleicht
Millionen. Wer es schafft, den unmittelbaren Ekel zu überwinden und die
daneben angebrachte Beschriftung zu lesen, der erfährt, daß dieses
Insektenheer ausschließlich aus den Nachkommen besteht, die ein einziges
Weibchen während seines Lebens hervorbringen kann — zumindest theoretisch.
Glücklicherweise wird diese potentielle Fruchtbarkeit nur selten in vollem
Umfang Wirklichkeit. Wie Darwin schon in der Entstehung der Arten feststellt,
könnten die meisten Lebewesen so viele Nachkommen hinterlassen, daß nicht
alle tatsächlich überleben können.
Irgend etwas wirkt diesem Potential
entgegen. (Wer keine Küchenschaben mag, wird dafür dankbar sein.) Zu diesem
Etwas gehören begrenztes Nährstoffangebot, Konkurrenten, natürliche Feinde,
widrige Klimabedingungen, Krankheiten und andere Einflüsse. Aber auch wenn
die durchschnittliche Individuenzahl über längere Zeit hinweg relativ stabil
bleibt, pendelt sie kurzfristig um den Mittelwert. Manchmal handelt es sich
dabei um geringe, manchmal aber auch um dramatische Schwankungen: Die
Population explodiert und bricht dann zusammen.
Wenn
man die Dynamik ökologischer Gemeinschaften in kurzen Zeiträumen — das
heißt über einige Jahrzehnte hinweg — verstehen will, ist es entscheidend,
daß man genau herausfindet, welche Kräfte für die Populationsschwankungen
der einzelnen Arten sorgen. Robert May betont: »Solche Kenntnisse sind nicht
nur von grundsätzlicher Bedeutung, sondern sie lassen sich auch praktisch
anwenden: Man kann versuchen, die Auswirkungen natürlicher oder vom Menschen
verursachter Veränderungen vorauszusagen, wie sie zum Beispiel eintreten,
wenn man eine Population dezimiert oder wenn sich die Klimabedingungen
ändern.«3
Stabilität
und Chaos in der Ökologie 189
Unter
der Überschrift »Gleichgewicht der Natur« sind die Populationsschwankungen
einfach zu erklären — zwar nicht im Detail, aber zumindest im Prinzip. Man
nimmt an, daß die Populationen der einzelnen Arten und die Gemeinschaften, zu
denen sie gehören, sich im Gleichgewicht oder in seiner Nähe befinden. Ohne
Störungen durch äußere Einflüsse erreichen die Wechselwirkungen zwischen
Pflanzen, Pflanzen- und Fleischfressern ein Fließgleichgewicht, in dem die
Populationen genau ausbalanciert sind. Das ist Fairfield Osborns
gleichmäßig brummende »koordinierte Maschine«.
Das
begrenzte Nahrungsangebot, die wechselseitige Konkurrenz, die Beute der
Räuber und sogar Krankheiten — all das gehört dazu, damit die Maschine
funktioniert. Hat eine Artengemeinschaft das Gleichgewicht erreicht, ist das
Klima der wichtigste störende Einfluß, entweder in Form langfristiger
Verschiebungen oder aber als plötzliche, heftige Episoden wie Stürme oder
Temperaturschwankungen. Klimaveränderungen begünstigen manche Arten und sind
für andere verheerend. Dezimiert beispielsweise ein Sturm die Population
einer bestimmten Pflanzenart, leiden auch die Pflanzenfresser, die von ihr
abhängig sind, und das wiederum vermindert das Nahrungsangebot für die
Raubtiere, die sich von den Pflanzenfressern ernähren, so daß die Population
zusammenbricht. Gleichzeitig haben andere Arten, die den Räubern ebenfalls
als Beute dienen, bessere Überlebenschancen, so daß deren Population
wächst. Ein einziger Sturm kann also dazu führen, daß die Populationen
einiger Arten explodieren, während andere zusammenbrechen.
Bei
solchen Populationsschwankungen geht das Gleichgewicht für einige Zeit
verloren. Die Individuenzahlen pendeln eine Zeitlang um den Mittelwert, aber
schließlich stellt sich die Balance wieder ein — bis zur nächsten
Störung. Da ökologische Gemeinschaften nur selten mit längeren Phasen ohne
äußere Erschütterungen gesegnet sind, schwanken Populationen die meiste
Zeit. Das klassische Beispiel in der ökologischen Fachliteratur ist die
Geschichte des kanadischen Luchses. Über 200 Jahre lang, von 1735 bis 1940,
fing man die Luchse wegen ihres Fells, und die Zahlen der von den Handels-
Gleichgewicht
der Natur? 190
firmen
aufgekauften Pelze waren für die Ökologen ein beispiellos gutes
Datenmaterial, um die Populationsentwicklung bei dieser Art
nachzuvollziehen. In den Befunden erkennt man eine deutliche
Gesetzmäßigkeit: Die Luchspopulation erlebte dramatische Phasen der Zu- und
Abnahme. Sie zeigte zum Beispiel zwischen 1830 und 1910 alle neun oder zehn
Jahre einen Höhepunkt, um dann schnell zusammenzubrechen. Das Muster
wiederholte sich ziemlich regelmäßig, aber die Zahl der Tiere an den
Höhepunkten schwankte beträchtlich, nämlich etwa zwischen 10.000 und
60.000.
Als
die Ökologen zum erstenmal diese Entwicklung analysierten, nahmen sie an, das
Muster sei die Folge der Räuber-Beute-Beziehung zwischen Luchsen und
Schneeschuhhasen, ihren wichtigsten Beutetieren. Wenn ein Räuber die
Population seiner Beutetiere stark dezimiert hat, nimmt sein Nahrungsangebot
drastisch ab, und dann wird auch die Population der Räuber kleiner. Wenn auf
diese Weise der Druck durch die Räuber nachläßt, kann sich die Population
der Beuteart erholen, und daraufhin vermehren sich auch die Räuber wieder.
Nach einer solchen einfachen Oszillation sah es auch bei kanadischen Luchsen
und Schneeschuhhasen aus.
Aber
wie sich herausstellte, war die Sache nicht so einfach. Die Hasenpopulation
ging wegen Unterschieden in ihrer Nahrungsversorgung auf und ab, und
die Luchspopulation machte offenbar die Schwankungen mit. Diese Vorstellung
erscheint durchaus logisch und verlängerte die Kette der Wechselwirkungen.
Aber der Verlauf war nicht vollkommen regelmäßig und sah an manchen Stellen
sogar völlig ungeordnet aus. Eine solche Kombination aus einer gewissen
Regelmäßigkeit und etwas scheinbar Zufälligem ist für die
Populationsschwankungen vieler Arten sogar geradezu typisch. Insektenplagen
folgen dieser Gesetzmäßigkeit ebenso wie die Populationsexplosionen bei den
Seeigeln im Nordatlantik und bei den Pazifischen Taschenkrebsen im
Nordwestpazifik. Wohin man in den ökologischen Gemeinschaften auch blickt,
erkennt man nicht nur Populationsschwankungen, sondern es sieht auch so aus,
als erfolgten die Veränderungen zufällig. Vom Meeresplankton bis zu den
Elefanten und von den Motten zu den Mäusen bietet sich das gleiche Bild.
191
|
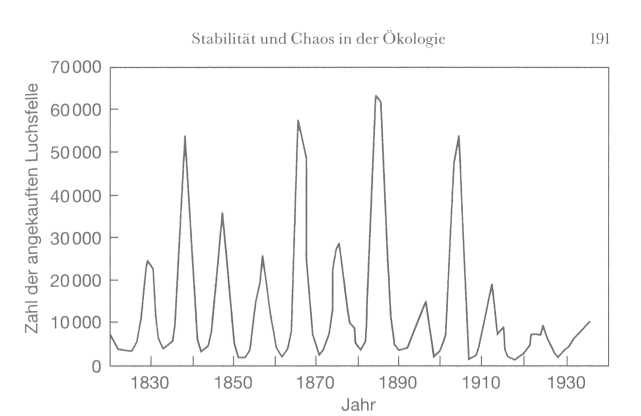 |
Das
klassische Beispiel für eine Art mit regelmäßig schwankender
Individuenzahl ist der Luchs. Früher glaubte man, die Ursache sei die
ungleichmäßige Wechselbeziehung zu den Schneeschuhhasen, den wichtigsten
Beutetieren der Luchse.
Wie man jedoch in jüngster Zeit erkannt hat, ist
die Ursache in Wirklichkeit die Beziehung zwischen den Hasen und ihren
Nahrungspflanzen; die Population der Luchse ist mehr oder weniger passiv vom
Bestand an Hasen abhängig.
|
Wie
läßt sich das unter dem Gesichtspunkt des natürlichen Gleichgewichts und
der Populationsbalance erklären? Die Antwort ist einfach: Alles, was man in
der Populationsentwicklung beobachtet — sei es nun Regelmäßigkeit,
scheinbare Zufälligkeit oder eine Kombination aus beidem —, gilt als
unmittelbare Folge äußerer Kräfte wie zum Beispiel
Klimaveränderungen. Die Tatsache, daß die Populationsentwicklung manchmal
unberechenbar ist, zeigt nur, wie komplex diese Störungen sind. So dachte man
zumindest.
Seit
etwa 20 Jahren wird diese Argumentation in Frage gestellt. Vielleicht, so die
Spekulation einiger Ökologen, läßt auch irgend etwas in der inneren Dynamik
der Lebensgemeinschaften die Entwicklungsmuster entstehen.
Gleichgewicht
der Natur? 192
Vielleicht
war die scheinbare Zufälligkeit kein Zufall, sondern ein Aspekt des
Phänomens, das man als Chaos bezeichnet. Wenn man hört, ein System sei
chaotisch, nimmt man meist an, es sei vom Zufall bestimmt und sein wirres
Verhalten habe keine einfache, analysierbare Grundlage. Aber wie jeder weiß,
der das Buch Chaos von James Gleick gelesen hat, kennt man in der
Mathematik seit einiger Zeit Systeme, die zwar wirr und nicht vorhersagbar,
aber nicht zufällig sind. Das Verhalten solcher Systeme läßt sich mit
mathematischen Gleichungen oft recht einfach beschreiben. Was dabei das
Paradoxe ist: Obwohl das System mathematischen Regeln unterliegt, kann sein
Verhalten höchst komplex und praktisch nicht vorhersagbar sein. Damit ist
grob und einfach gesagt, was die Mathematiker unter deterministischem Chaos
verstehen.
Chaotische
Systeme kennt man heute in vielen Bereichen der Physik, so bei der Entstehung
des Wetters und bei Turbulenzen in Flüssigkeitsströmungen. Kaum jemandem ist
aber klar, daß Populationsschwankungen in ökologischen Gemeinschaften zu
den ersten Phänomenen gehörten, die man als möglichen Ausdruck chaotischen
Verhaltens untersuchte. Genau das tat Robert May vor 20 Jahren, und dann
beschrieb er seine Arbeiten in der Fachzeitschrift Nature in einem
Aufsatz, der zum Klassiker wurde. Die Biologen wagten sich nur langsam auf den
Weg, den May aufgezeigt hatte, vor allem weil sie an der Vorstellung vom
Gleichgewicht der Natur und den ausbalancierten Populationen hingen, zum Teil
aber auch weil derartige biologische Fragestellungen weitaus komplexer und
schwieriger zu analysieren sind als jedes physikalische System. May schrieb
einmal: »Für manche Ökologen hat das [Chaos] ein Flair von schwarzer
Magie.«4
Besessen von der Vorstellung vom Gleichgewicht, suchten die Ökologen
weiterhin nach Argumenten, die dafür sprachen, und ignorierten mit schöner
Regelmäßigkeit das unberechenbare Verhalten, das daraufhinwies, daß sich in
Wirklichkeit etwas anderes abspielte.
In
den letzten ein oder zwei Jahren hat man aber in Freilandexperimenten und
theoretischen Modellen die lange gesuchten Indizien für echtes chaotisches
Verhalten in ökologischen Gemeinschaften entdeckt.
193
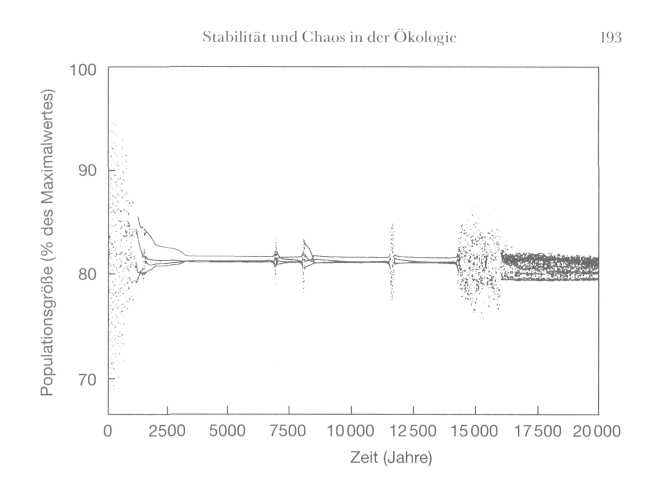
Computersimulation
für die Entwicklung einer Population Pazifischer Taschenkrebse.
Gelegentlich schwankt der Bestand stark, auch wenn es keinen äußeren
Auslöser gibt. Hier wird deutlich, welche unerwarteten und
unberechenbaren Auswirkungen das Chaos in lebenden Systemen haben kann.
(Mit freundllicher Genehmigung von Alan Hastings und Kevin Higgins.)
Heute
sind wir gezwungen, die Natur und die formenden Kräfte für das, was wir
sehen und erleben, unter einem anderen Blickwinkel zu betrachten. Das
widerspricht zutiefst unserem Gefühl und ist deshalb nur schwer zu
akzeptieren.
Mitte
der achtziger Jahre stellte David Tilman, ein Ökologe der University of
Minnesota, die Frage: Wie beeinflussen unterschiedliche
Stickstoffkonzentrationen im Boden das Wachstum einer amerikanischen
Wildgrasart? Als er das Experiment plante, ging es ihm nicht um chaotische
Vorgänge, aber er war so aufgeschlossen, daß er sie erkannte, als er sie
sah. Bei geringem Stickstoffgehalt im Boden war das Wachstum über fünf Jahre
hinweg konstant, unabhängig davon, ob er die Samen reichlich oder sparsam
ausstreute.
Gleichgewicht
der Natur? 194
Bei
hoher Stickstoffkonzentration ergab sich jedoch ein ganz anderes Bild, zu dem
auch das klassische Anzeichen für chaotisches Verhalten gehörte: wildes,
unberechenbares Auf und Ab. Einmal schrumpfte die Graspopulation auf ein
Sechstausendstel ihrer anfänglichen Größe, das heißt fast bis auf Null.
Die Beschreibung der Vorgänge, die sich auf dem Feld tatsächlich abspielten,
hört sich banal an: Der viele Stickstoff im Boden sorgte für schnelles,
üppiges Wachstum; im Winter starben die Pflanzen ab und bedeckten den Boden
mit einer dicken Abfallschicht, die im folgenden Frühjahr das Wachstum
verhinderte; so kam es zu einem Auf und Ab von wechselnder Stärke. Mäßiges
Wachstum, angetrieben von mäßigem Stickstoffgehalt, führte zu einer
stetigeren Populationsentwicklung. Als Tilman seine Befunde Ende 1991
veröffentlichte, war die Reaktion eine Mischung aus Verblüffung und
Begeisterung.
Dieser
Bereich der Ökologie ist stark von theoretischen Arbeiten geprägt, nicht
zuletzt weil Experimente, wie Tilman sie durchführte, in Planung und
Ausführung nicht einfach sind. Wie ich bereits erwähnt habe, ist die
experimentelle Handhabung von Ökosystemen von berüchtigter Schwierigkeit. Im
gleichen Jahr, als Tilman seine Befunde bekanntgab, veröffentlichten Robert
May und zwei seiner Kollegen die Ergebnisse eines mathematischen Modells mit
einem parasitenartigen Organismus und seinem Wirt, die sich ganz ähnlich
verhielten wie Tilmans System. Die Berechnungen zeigten unberechenbare
Veränderungen der Population im Verlaufe vieler »Generationen«,
hervorgerufen ohne äußere Störungen allein durch die Wechselwirkungen
zwischen den »Arten«. Das Modell beschrieb die Arten und ihre Beziehungen in
mathematischen Gleichungen. Die komplexe Dynamik des Systems hatte ihren
Ursprung im System selbst und wurde ihm nicht von außen aufgezwungen. Ebenso
wichtig ist in diesem Fall, daß das scheinbar zufällige, unberechenbare
Verhalten sich aus einfachen mathematischen Beziehungen ergab — ein
Anzeichen für echtes Chaos. Etwas Ähnliches beobachteten Alan Hastings und
Kevin Higgins von der University of California in Davis später auch an ihrem
Modell der Population von Pazifischen Taschenkrebsen.
Stabilität
und Chaos in der Ökologie 195
Sie
beschrieben — wiederum mit einfachen mathematischen Gleichungen — die
Spezies und ihr Verhalten entlang eines theoretischen Küstenabschnitts.
Wieder zeigte sich ein zufälliges, unberechenbares Verhalten mit Phasen der
Stabilität und Zeiten heftiger Populationsschwankungen. »Bevölkerungsexplosionen ohne jede Veränderung der biologischen oder
physikalischen Bedingungen dürften eine grundlegende Eigenschaft dieser
Dynamik sein«, schrieben sie Anfang 1994 in ihrer Veröffentlichung in der
Fachzeitschrift Science.5
Der
Nachweis, daß Populationsgrößen durch Wechselwirkungen innerhalb des
Systems und ohne äußere Veränderungen drastisch und unvorhersehbar
schwanken können, war ein wichtiger Schritt zur Aufklärung der
Gesetzmäßigkeiten, die wir in der Natur beobachten. »Die Vorstellung vom
Chaos ist amüsant und gleichzeitig ein wenig bedrohlich«, meinen William
Schaffer und Mark Kot, zwei Ökologen der University of Arizona, die viel dazu
beigetragen haben, die Kenntnisse über das Chaos in Ökosystemen zu
erweitern. »Einerseits bietet sie eine deterministische Alternative zu der
Idee, Populationsschwankungen seien ausschließlich die Folge äußerer
Störungen. Gleichzeitig könnte die Chaostheorie aber auch das
Begriffsgebäude der heutigen Ökologie ins Wanken bringen.«6
Die
ökologischen Vorstellungen werden durch solche Erkenntnisse mit Sicherheit
erschüttert, aber unter dem Gesichtspunkt der biologischen Vielfalt ist Chaos
eine positive Kraft. Wie ich bereits erklärt habe, können in stabilen
Populationen wenige Arten oder auch nur eine einzige die Oberhand gewinnen.
Durch Populationsschwankungen hingegen können die Lebensgemeinschaften zu
einem größeren Artenreichtum gelangen. Wie man daran erkennt, trägt das
scheinbar zufällige Verhalten, das sich aus der inneren Dynamik der
ökologischen Gemeinschaften ergibt, zur Verstärkung der Vielfalt bei.
Das
war eine unerwartete Entdeckung, aber offenbar hält die Chaostheorie für die
Ökologen noch mehr Überraschungen bereit. Zu Beginn dieses Kapitels habe ich
die mosaikartige Struktur vieler Lebensräume beschrieben, wobei die
Mosaiksteine ähnliche, aber unterschiedliche ökologische Gemeinschaften
darstellen.
Gleichgewicht
der Natur? 196
Nach
der herkömmlichen Vorstellung spiegeln sich in diesen Unterschieden kleine,
aber wichtige Abweichungen der physikalischen Umweltbedingungen wider. Die
Chaostheorie bietet eine andere Erklärung. Wie Robert May und seine Kollegen
mit ihrem Modell von Parasiten und Wirt entdeckten, schwankt im Laufe der Zeit
nicht nur die Größe der Population, sondern sie kann auch im Raum
ungleichmäßig — mosaikartig — verteilt sein. Sie arbeiteten mit Modellen, in
denen drei oder mehr Arten in einer theoretischen Landschaft verteilt waren,
und stellten dabei fest, daß die Arten durch die Dynamik der Wechselwirkungen
oft getrennt bleiben. »Im Extremfall führte das ... zur Entstehung kleiner,
relativ unbeweglicher >Inseln< innerhalb des Lebensraumes, die wie
isolierte, begünstigte Bereiche aussahen«, schrieben sie in ihrem
Fachartikel, der im Sommer 1994 in Nature erschien.7
Schwankungen
in der Verteilung der Arten innerhalb der Lebensräume (beispielsweise der
Bäume im Wald oder der Blumen auf den Wiesen) kommen häufig vor und wurden
mit Unterschieden in Konkurrenz- und Ausbreitungsfähigkeit erklärt, das
heißt als Teil der Reaktion auf die Verteilung der physikalischen Bedingungen
in dem Lebensraum. Aus der Sicht der Chaostheorie, die der Intuition
widerspricht, reicht diese Erklärung nicht aus. Die mosaikartige
Zusammensetzung, die man in der Natur beobachtet, kann sich aus der inneren
Dynamik des Ökosystems auch dann ergeben, wenn das Umfeld der verschiedenen
Gemeinschaften genau gleich ist.
Wir
sehen also, daß die Natur sich keineswegs im Gleichgewicht befindet; sie ist
keine »koordinierte Maschine«, die stets nach Ausgleich strebt. Das Ganze
ist wesentlich interessanter. Zwar ist nicht zu leugnen, daß die Anpassung an
örtliche physikalische Gegebenheiten und an äußere Kräfte wie das Klima
dazu beitragen, der Welt ihre Gestalt zu geben. Aber ebenso offenkundig ist
auch, daß die beobachteten räumlichen und zeitlichen Verteilungsmuster zu
einem großen Teil aus der Natur selbst erwachsen. Das ist eine aufregende
Erkenntnis, aber sie bedeutet auch, daß die Arbeit der Naturschützer noch
schwieriger wird.
Stabilität
und Chaos in der Ökologie 197
Früher
glaubte man, die Populationsgröße lasse sich steuern, indem man die
äußeren Bedingungen beeinflußte (soweit das möglich war). Heute müssen
wir erkennen, daß diese Möglichkeit nicht so plausibel ist, wie es früher
schien. Die Natur in ihrer unendlichen Vielfalt und mit ihren unendlich
komplizierten Abläufen zu verstehen und dann zu akzeptieren und
einzugestehen, daß alle Versuche, sie zu lenken, so gut wie nutzlos sind, ist
viel besser, als sich aus Unkenntnis vorzustellen, ein solcher Einfluß sei
möglich.
Im
Jahr 1789 brachte Gilbert White, ein Geistlicher in Südengland, ein
wunderhübsches kleines Buch mit dem Titel The Natural History of Selborne (Die
Naturgeschichte von Seiborne) heraus. Der bescheidene Band, der auf der Liste
der am häufigsten nachgedruckten englischsprachigen Bücher an vierter Stelle
steht, enthält eine Sammlung scharfsichtiger Naturbeobachtungen aus dem Dorf,
in dem White als Seelsorger lebte, und seiner Umgebung. Natürlich bediente er
sich nicht der Sprache der heutigen Ökologie; er sprach nicht von
Lebensgemeinschaften, Nahrungsnetzen und trophischen Ebenen. Aber er erkannte
die Wechselwirkungen zwischen den Arten und die unterschiedlichen Ebenen ihrer
gegenseitigen Abhängigkeit, und damit war er bei einer entscheidenden Frage,
die auch heute die Ökologen beschäftigt wie keine andere: Was bestimmt
darüber, daß eine Lebensgemeinschaft so wird, wie sie ist? Genauer läßt
sie sich in eine grobe Alternative fassen: Geschieht es durch Planung oder
durch Zufall? Gezielter kann man fragen: Haben die Arten in einer bestimmten
Lebensgemeinschaft etwas Besonderes, so daß diese und nur diese
Artenzusammensetzung für den Lebensraum optimal ist? Mit anderen Worten: Was
für eine Ordnung liegt Darwins »Fleckchen Boden« zugrunde?
Durch
Betrachten der wirklichen Welt läßt sich diese Frage nicht ohne weiteres
beantworten, denn der zeitliche und räumliche Umfang der Ökosysteme entzieht
sich jeder einfachen Analyse. Deshalb spielen sich die bahnbrechenden Arbeiten
der Ökologie derzeit zu einem großen Teil in Computern ab, in denen die
Wissenschaftler experimentelle Ökosysteme simulieren. Im Vergleich zur Natur
sind solche Systeme sehr einfach, aber in den letzten Jahren haben sie
neue Erkenntnisse geliefert, die wie der Folgerungen der Chaostheorie
entschieden der Intuition widersprechen.
Gleichgewicht
der Natur? 198
Vor
etwa zehn Jahren bauten beispielsweise Stuart Pimm und Mac Post an der
University of Tennessce ein solches Ökosystem zusammen, zu dem sie jeweils
eine Art nach der anderen — Pflanzen, Pflanzenfresser und Fleischfresser —
hinzufügten. Jede Art wurde mathematisch mit einer Sammlung von
Verhaltensweisen beschrieben, zu denen Typ und Größe sowie Platz- und
Nahrungsbedarf gehörten. Damit vollzogen Pimm und Post im Computer nach, was
in der Natur bei der Besiedelung jungfräulicher Gebiete geschieht,
beispielsweise nach einem Waldbrand oder auf einer neuentstandenen
Vulkaninsel. Eine ökologische Gemeinschaft findet sich langsam zusammen; den
Vorgang nennt man Sukzession: Er beginnt mit den einfachsten Lebewesen, die in
einem verödeten Lebensraum gedeihen können, und allmählich kommen immer
mehr Arten hinzu, die auf die bereits vorhandenen angewiesen sind.
Pflanzenfresser können zum Beispiel nicht dazugehören, solange sich keine
Pflanzen angesiedelt haben, und Raubtiere sind erst möglich, wenn die
Beutetiere vorhanden sind.
In
dem Computermodell kamen neue Arten nach dem Zufallsprinzip zu der
Lebensgemeinschaft hinzu. Es gab keine Versuche, dem System eine bestimmte
Zusammensetzung zu verleihen, sondern es sollte sich von selbst entwickeln.
Und wie in der Natur mußten die Pflanzen vor den Pflanzenfressern und diese
vor den Fleischfressern vorhanden sein. Die Lebensgemeinschaft baute sich nach
einer verblüffenden Dynamik auf. Anfangs konnte man leicht neue Arten
hinzufügen (vorausgesetzt, sie waren ökologisch plausibel) . Aber je mehr
die Gemeinschaft wuchs (das heißt je mehr Arten sie umfaßte), desto
schwieriger wurde es für neue Arten, sich einzufügen. Als das Ökosystem aus
etwa zwölf Arten bestand, wurde die Einwanderung recht schwierig, und wenn
sie gelang, führte sie oft zum Verlust einer oder mehrerer der zuvor
vorhandenen Arten. Das erinnerte an Darwins Vergleich mit den Keilen, wonach
die Arten dicht gedrängt sind, so daß jedesmal, wenn man einen neuen Keil
hineintreibt, ein anderer herausgedrückt wird.
Stabilität
und Chaos in der Ökologie 199
Ökologisch
ausgedrückt, beobachteten Pimm und Post den Erfolg fremder Arten, die in eine
Gemeinschaft einwanderten, und ihre Auswirkungen auf diese Gemeinschaft. In
artenarmen Gemeinschaften gelang die Einwanderung leicht, bei hoher Artenzahl
war sie jedoch schwierig. Der britische Ökologe Charles Elton hatte schon vor
über 30 Jahren die Vermutung geäußert, daß es sich in der Natur genauso
verhält. Aber warum?
Nach
der herkömmlichen Erklärung werden mit zunehmender Artenzahl in einer
Gemeinschaft immer mehr ökologische Nischen besetzt. Ein potentieller
Einwanderer, der eine bereits besetzte Nische vorfindet, hat es schwerer, sich
durchzusetzen, als wenn die Nische noch frei ist. Im ersten Fall muß der
Eindringling die vorhandene Art im Konkurrenzkampf verdrängen, um zu einem
Teil der Gemeinschaft zu werden; im zweiten ist das nicht notwendig. Das
erscheint ökologisch plausibel, ist aber offensichtlich falsch. Der
potentielle Einwanderer muß sich nicht mit der Art auseinandersetzen, die
seine bevorzugte Nische besetzt hält, sondern mit der Gemeinschaft als
Ganzem. Sehr deutlich wurde das in Computermodellen, die Ted Case von der
University of California in San Diego konstruierte. Er simulierte mehrere
unterschiedliche Lebensgemeinschaften und änderte jeweils den Umfang der
Wechselbeziehungen zwischen den beteiligten Arten: Die Interaktionen waren
einmal stark, ein anderes Mal schwach.
»Gemeinschaften
mit vielen, stark in Wechselbeziehung stehenden Arten schränken die
Einwanderungsmöglichkeiten für die meisten Arten ein«, schrieb Case über
seine Befunde. »Solche Gemeinschaften richten selbst für einen überlegenen,
neu hinzukommenden Konkurrenten eine Art <Aktivierungsschranke> auf, die
Eindringlinge abhält, wenn sie in geringer Individuenzahl auftauchen.«8
Wenn
die Nischenhypothese stimmt, sollte man erwarten, daß ein potentieller
Eindringling Erfolg hat, wenn er der vorhandenen Art im Konkurrenzkampf
überlegen ist. Aber das ist nicht der Fall. Gemeinschaften, deren einzelne
Arten in enger Wechselbeziehung stehen, sind für das Eindringen neuer Arten
selbst dann weniger anfällig, wenn es sich dabei um überlegene Konkurrenten
handelt.
Gleichgewicht
der Natur? 200
»Diese
Modelle weisen daraufhin, daß der unterschiedliche Erfolg der Eindringlinge
weniger von ihren Eigenschaften als von den Eigenschaften der Gemeinschaft
abhängt«, schließt Case.9 Wenn dieser Befund stimmt, ist er von größter
Bedeutung nicht nur für tiefgreifendere Kenntnisse über die Ökosysteme,
sondern auch für den Naturschutz. Häufig versucht man, in einem Ökosystem
eine Art zu schützen, die einer einwandernden, exotischen Art unterlegen ist.
Wie Cases Modelle zeigen, hat ein solcher unterlegener Konkurrent die besten
Überlebenschancen, wenn er zu einer artenreichen, intakten und ungestörten
Lebensgemeinschaft gehört. Wenn man eine Gemeinschaft als Ganzes vor
Störungen schützt, bietet man also ihren schwächsten Mitgliedern
Sicherheit, weil man ein schützendes Netzwerk schafft.
Der
Ausdruck »schützendes Netzwerk« hat etwas unverkennbar Mystisches, und
deshalb müssen wir verstehen, was damit gemeint ist. Er bezeichnet die
Nahrungsnetze, nach einer Erklärung »die Straßenkarten für Darwins
Stückchen Erde... die zeigen, wie eine Gemeinschaft sich zusammenfindet und
wie sie funktioniert«.10 Die Karten zeigen die Wechselwirkungen zwischen den
Arten in einer Gemeinschaft und geben beispielsweise Auskunft darüber, wer
wen frißt. Die Nahrungsnetze sehen oft verwirrend kompliziert aus, und
anfangs glaubte man, jede Gemeinschaft habe ihr eigenes, einzigartiges
Nahrungsnetz. Als die Biologen aber die vordergründige Komplexität
durchdrungen hatten, erkannten sie, daß alle Nahrungsnetze sich unabhängig
von der Art der Lebensgemeinschaft sehr ähnlich sind und einige gemeinsame
Eigenschaften haben. Dazu gehören zum Beispiel die Länge der Nahrungsketten,
die etwas darüber aussagen, wer wen frißt, und das Zahlenverhältnis von
Räubern und Beutetieren. Wohin man in der Natur auch blickt, erkennt man
ähnliche Gesetzmäßigkeiten. Die Tatsache, daß man solche Gemeinsamkeiten
sieht, obwohl eine unbegrenzte Vielfalt möglich wäre, sagt etwas
Grundlegendes über die der Natur zugrundeliegende Ordnung aus. Diese Ordnung
erwächst offensichtlich aus der inneren Dynamik des Systems selbst und wird
ihm nicht durch äußere Umstände aufgezwungen.
Stabilität
und Chaos in der Ökologie 201
Die
Wechselwirkungen, die Ted Gase in seinem Computer manipulierte, stellten die
Nahrungsnetze der wirklichen Welt dar. Das schützende Netzwerk, das er in den
Gemeinschaften mit engen Wechselbeziehungen erkannte, läßt sich also als
Eigenschaft der zugrundeliegenden Nahrungsnetze erklären. Obwohl seine
Beobachtung der Intuition widerspricht, braucht man keine mystische Kraft
heranzuziehen, um sie zu erklären.
Die
Ökosysteme, die sich in den Computermodellen von Stuart Pimm und Mac Post
zusammenfanden, zeigten ein Geflecht von Wechselwirkungen zwischen den Arten,
das stark den Nahrungsnetzen der wirklichen Ökosysteme ähnelte. Das ließ
die Überzeugung wachsen, daß diese Modelle trotz ihrer Einfachheit
realistisch waren. Wie bereits erwähnt, lautete das erste Ergebnis: In
artenarme Gemeinschaften können neue Arten leicht einwandern, bei Systemen
mit vielen Arten ist das erheblich schwieriger. Schwieriger, aber nicht
unmöglich. Eine artenreiche Gemeinschaft, die man heranreifen läßt, bleibt
nicht unverändert, sondern es kommt zu einer langsamen Artenfluktuation. Mit
anderen Worten: Manche neu hinzukommenden Arten setzen sich durch, in der
Regel indem sie andere verdrängen; die Zusammensetzung der Gemeinschaft ist
nicht statisch, sondern dynamisch. Eine erfolgreiche Art kann später
ebenfalls als Opfer eines neuen Eindringlings verdrängt werden, aber dann
hinterläßt sie in der Gemeinschaft ihre Spuren wie einen Fußabdruck im
Sand. So gelangten Pimm und Post zu ihrem zweiten Ergebnis: Bei reifen, artenreichen
Gemeinschaften ist das Eindringen viel schwieriger als bei neu entstandenen.
Offenbar stärkt irgendein Element des Reifungsprozesses das schützende
Netzwerk innerhalb der Gemeinschaft. Die Gemeinschaft scheint sich weiterzuentwickeln — sie wird scheinbar fast zielgerichtet besser, und zwar
auf eine Art, die nur schwer einzugrenzen ist.
Das
Ergebnis ist keineswegs die Laune eines weltfremden Computermodells ; in der
Natur geschieht genau das gleiche. Ein Beispiel ist Hawaii. Dort gibt es zwei
Arten von Ökosystemen: erstens die Bergwälder, die nicht durch menschliche
Eingriffe gestört wurden und
Gleichgewicht
der Natur? 202
eine
ausgereifte, artenreiche Lebensgemeinschaft bilden; und zweitens die Wälder
der Niederungen, die einschließlich ihrer Umgebung durch die Tätigkeit der
Menschen verändert wurden. Wenn sie sich von solchen Störungen erholen,
befinden sie sich, obwohl sie artenreich sind, in einem unreifen
Aufbaustadium. Seit vor 1500 Jahren die ersten polynesischen Siedler
eintrafen, wurden die Inseln viele Male kolonisiert, und mit den
Neuankömmlingen kamen auch viele neue Arten, die entweder absichtlich
mitgebracht oder zufällig eingeschleppt wurden. So wurden auf Hawaii
beispielsweise mehr neue Vogel- und Pflanzenarten eingeführt als irgendwo
sonst auf der Welt. 28 Prozent der Insekten- und 65 Prozent der Pflanzenarten
des Archipels sind ursprünglich nicht hier zu Hause. Auch sämtliche
Säugetiere kamen erst in jüngerer Zeit hierher.
Vor
30 Jahren schrieb Charles Elton in seinem Buch The Ecology of Invasions by
Animals and Plants, es handele sich um »eine der großen
Vermischungen in der Tier- und Pflanzenwelt der Erde«. Jedesmal wenn eine
fremde Art sich durchsetzen konnte, führte das zu einer Schrumpfung der
Populationen einheimischer Arten, oder es trieb sie in ganzen Wellen zum
Aussterben. Die Frage ist nun: Wo setzten die fremden Arten sich durch? In den
unreifen Ökosystemen der Niederungen oder in den alten Lebensgemeinschaften
der Bergwälder? Die Antwort: in ihrer großen Mehrzahl in den ersteren. Die
ausgereiften Ökosysteme konnten der Invasion offenbar besser widerstehen als
die unreifen. Oder in der Sprache der theoretischen Ökologie: Die reifen
Ökosysteme hatten einen dauerhaften Zustand erreicht.
Reife
Lebensgemeinschaften — ob in der wirklichen Welt oder im Computer — haben
ganz offensichtlich wichtige ökologische Eigenschaften, die unreifen Systemen
fehlen. Und daraus läßt sich der naheliegende Schluß ziehen, daß es
während des Aufbaus zu einer Selektion von Arten kommt, die in irgendeiner
Form überlegen sind. Pflanzenarten könnten zum Beispiel in der
Produktivität überlegen sein, Räuber in Schnelligkeit oder Tarnung, und so
weiter. Und natürlich ist eine Gemeinschaft überlegener Arten ökologisch
gegenüber einer solchen aus unterlegenen Arten im Vorteil.
Stabilität
und Chaos in der Ökologie 203
Als
Pimm und Post aber in ihrem Computermodell die Verhaltensmerkmale der
Arten in den dauerhaften Gemeinschaften untersuchten, konnten sie keine
Anzeichen von Überlegenheit finden. Oder ökologisch ausgedrückt: Diese
Arten unterschieden sich nicht von solchen, denen es nicht gelungen war, sich
in der Gemeinschaft zu etablieren. Vielleicht, so die Spekulation der
Wissenschaftler, hatten sie nicht die richtigen Eigenschaften betrachtet.
Wie
sich herausstellte, hatten sie keinen Fehler gemacht. Dies wurde deutlich, als
Jim Drake, ein anderer Ökologe, der damals an der Purdue University
arbeitete, ähnliche Computersimulationen ablaufen ließ. Wie Pimm und Post,
so sorgte auch Drake für den Aufbau einer ökologischen Gemeinschaft, indem
er eine Art nach der anderen hinzufügte. Neu war aber, daß er dabei aus
einem begrenzten Spektrum von insgesamt 125 Arten schöpfte. Gelang einer
davon das Eindringen nicht, stand sie später für einen weiteren Versuch zur
Verfügung. Wieder entwickelte sich eine dauerhafte Gemeinschaft mit etwa
einem Dutzend Arten. Dann begann Drake mit dem gleichen Artenspektrum von
vorn, und der Vorgang der Reifung bis zu einer dauerhaften Gemeinschaft mit
einem Dutzend Arten wiederholte sich. Aber es war eine andere Gemeinschaft,
die noch nicht einmal die Hälfte ihrer Arten mit der ersten gemeinsam hatte.
Er ließ den Vorgang noch einige Dutzend Male ablaufen: Jedesmal erhielt er
eine reife, dauerhafte Gemeinschaft, aber jedesmal mit einer anderen
Artenzusammensetzung. Auch hier war keine der Arten in den Gemeinschaften in
irgendeiner Hinsicht erkennbar »besser« als die anderen. Jede Art konnte zum
Mitglied einer dauerhaften Gemeinschaft werden, wenn sie zum richtigen
Zeitpunkt hinzukam.
Diese
Befunde sind gleichermaßen faszinierend und wichtig. Zunächst einmal sieht
man daran, daß dauerhafte Gemeinschaften sich aus zufällig hinzukommenden
Arten bilden können. Und zweitens erwächst die ökologisch entscheidende
Eigenschaft der Beständigkeit oder Stabilität aus den Wechselbeziehungen der
Arten in der Gemeinschaft und nicht aus den überlegenen Eigenschaften der
einzelnen Arten. Mindestens ebenso bedeutsam sind die Folgerungen, die sich
daraus für die mosaikartige Zusammensetzung der Natur ergeben.
204
Wie
wir bereits gesehen haben, erklärt man die Unterschiede zwischen benachbarten
Ökosystemen in der Regel mit den unterschiedlichen physikalischen
Bedingungen. Außerdem haben wir erfahren, daß man nach der Chaostheorie mit
einem solchen Flickenmuster auch dann rechnet, wenn es in der Umwelt keine
physikalischen Unterschiede gibt. Die Arbeiten von Pimm, Post und Drake zeigen
eine weitere Ursache der mosaikartigen Zusammensetzung auf, die ebenfalls
nichts mit der Anpassung an die lokale Umwelt zu tun hat: die Geschichte. Die
endgültige Zusammensetzung eines dauerhaften Ökosystems hängt eindeutig
davon ab, in welcher Reihenfolge die einzelnen Arten während der Reifung des
Systems hinzugekommen sind. Manchmal ist es für eine Art von Vorteil, wenn
sie früh auf der Bildfläche erscheint, ein anderes Mal ist es besser, wenn
sie spät dran ist. Alles hängt davon ab, welche Arten bereits zu der
Gemeinschaft gehören. Wie wir in einem früheren Kapitel gesehen haben,
erkennt man in der Geschichte oder im Zufall immer stärker eine wichtige
Kraft, die den Lauf der Evolution mitgestaltet, und der Anpassung gesteht man
heute weniger Bedeutung zu als früher. Analog sind die Verhältnisse auch
hier: Die Geschichte trägt machtvoll dazu bei, die Evolution der Ökosysteme
zu gestalten, und die Anpassung spielt eine eher untergeordnete Rolle. Diese
Sichtweise für die Natur unterscheidet sich stark von der bisherigen
Vorstellung.
Wenn
sie auch für die wirkliche Natur gilt. Jim Drake stellte sie auf den
experimentellen Prüfstand, indem er mit Mikroorganismen (vor allem Algen
verschiedener Arten) das gleiche tat wie mit den Arten im Computer. Wenn er
Arten nach dem Zufallsprinzip hinzufügte, erhielt er viele verschiedene
dauerhafte Gemeinschaften. Die Geschichte ist tatsächlich von Bedeutung.
Kürzlich betrachteten zwei Paläontologen das Ganze unter dem Gesichtspunkt
der Fossilfunde. Martin Buzas von der Smithsonian Institution und Stephen
Culver vom Londoner Natural History Museum untersuchten die Zusammensetzung
küstennaher Gemeinschaften von Meeresbewohnern an der nordamerikanischen
Atlantikküste über 55 Millionen Jahre hinweg. In dieser Zeit stieg und fiel
der Meeresspiegel sechsmal.
Stabilität
und Chaos in der Ökologie 205
Sechsmal
bildeten sich in dem küstennahen Lebensraum neue Gemeinschaften, deren
Mitglieder aus dem Artenspektrum der Umgebung stammten. Und sechsmal war die
Zusammensetzung der Gemeinschaften anders. In einem Kommentar zu den Befunden
schrieb Jeremy Jackson, ein Ökologe der Smithsonian Institution: »Dies ist
sicher der Todesstoß für die Vorstellung von eng verzahnten ökologischen
Gemeinschaften im Meer.« Das stimmt, und gleichzeitig ist es ein Beleg für
die Bedeutung historischer Zufälle.
Wenn
schon diese Ergebnisse der Intuition zu widersprechen scheinen, so gilt das im
doppelten Sinn für eine weitere Beobachtung von Jim Drake. Er sagte sich
sinngemäß folgendes: »Die dauerhaften Gemeinschaften, die ich in meinem
Computer aufgebaut habe, funktionieren ganz offensichtlich sehr gut. Ich werde
also eine davon nehmen und sie noch einmal ganz von vorn aufbauen, und dazu
benutze ich nur das ungefähre Dutzend Arten, das auch jetzt zu der
Gemeinschaft gehört.« Es gelang ihm nicht. Nachdem er die Gemeinschaft
auseinandergenommen hatte, konnte er sie nicht wieder zusammensetzen, ganz
gleich, in welcher Reihenfolge er die Arten hinzufügte.
Stuart
Pimm bezeichnet es als »Holperstreckeneffekt«, und das mit gutem Grund. Die
Erklärung ist ein wenig abgehoben-mathematisch, aber im wesentlichen besagt
sie folgendes: Um den dauerhaften Zustand Z zu erreichen, muß das Ökosystem
die Stadien A bis Y durchmachen. Man kann nicht in einem Schritt nach Z
gelangen. Heute interessiert man sich immer stärker für die
Wiederherstellung verarmter oder zerstörter Ökosysteme — zwei Beispiele
sind die Prärien des amerikanischen Mittelwestens und die Everglades-Sümpfe
in Florida.
In
solchen Fällen kennt man aus historischen Dokumenten häufig die
ursprüngliche Artenzusammensetzung. Bevor man zu den gerade beschriebenen
Erkenntnissen gelangt war (die immer noch weiter verfeinert werden), neigten
die Ökologen dazu, einfach die Arten für das Ökosystem, das man
wiederherstellen wollte, zusammenzubringen und in dem gewählten Lebensraum
sich selbst zu überlassen. Mehrmals standen sie dann vor einem Rätsel, wenn
es nicht funktionierte. Jetzt wissen wir, warum.
206
Wir
haben gesehen, daß die Natur nicht immer so ist, wie sie zu sein scheint. In
den ökologischen Gemeinschaften gibt es eine Dynamik, die der Intuition
widerspricht und mit der man deshalb nicht gerechnet hatte.
Lebensgemeinschaften wandeln sich ständig und verbessern sich scheinbar
zielgerichtet, aber mittlerweile wissen wir, daß Zufall und Geschichte dabei
eine große Rolle spielen. Ich möchte dieses Kapitel mit einem Bericht über
ein reales Ökosystem beschließen; er läßt die Dynamik erkennen, zeigt die
Bedeutung des Zufalls und ist eine heilsame Geschichte für
Möchtegern-Naturschützer.
Im
Norden von Botswana liegt der Chobe-Nationalpark, ein typisches Beispiel für
eine ganze Reihe von Ökosystemen in Süd- und Ostafrika. Hier gibt es
zahlreiche große Pflanzenfresser, von denen manche auch Wanderungen machen,
darunter Giraffen, Büffel, Elefanten, Zebras, Gnus und
Schwarzfersenantilopen. Die ebenfalls reichhaltige Bevölkerung der
Fleischfresser besteht aus Löwen, Hyänen, Wildhunden und Schakalen. Der
mosaikartige Lebensraum aus Graslandschaften und Akaziengehölzen beherbergt
vielfältige Vogel- und Insektenarten. Insgesamt bietet der Park eine reiche
Artenfülle, wie man sie sich unter dem Begriff »Wildnis« vorstellt.
Die
Leitung des Parks möchte diese Vielfalt erhalten, denn sie ist für Touristen
attraktiv und wirkt, als sei alles in Ordnung. In Wirklichkeit gibt es aber
ein großes Problem: Die Akaziengehölze werden zerstört, vor allem von
Elefanten, und es wachsen keine neuen Bäume nach. Wenn die bewaldeten Gebiete
auf einen Bruchteil ihres jetzigen Umfanges schrumpfen, werden die
Parkverwalter glauben, sie hätten versagt, denn sie wollen, daß alles so
bleibt, wie es ist. Aber das zu erreichen, wäre nicht nur ökologisch falsch,
sondern wahrscheinlich auch unmöglich. Ein Blick auf die ökologische
Vergangenheit des Parks zeigt, warum.
Das
wichtigste Oberflächengewässer der Region ist der Savuti-Kanal. Wenn er
gefüllt ist, fließt er von Angola über die Linyanti-Sümpfe und mündet in
die Savuti-Marsch (die derzeit eine Graslandschaft ist). Er war im 19.
Jahrhundert gefüllt, trocknete um die Jahrhundertwende aus und blieb bis
Mitte der fünfziger Jahre trocken.
207
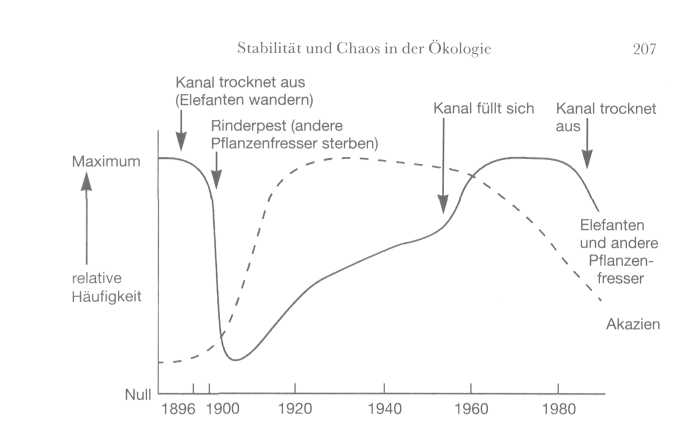
Die
meisten Lebensräume machen zyklische Veränderungen durch, die von
inneren und äußeren Kräften angetrieben werden.
Das Diagramm zeigt die
Entwicklung im Chobe-Nationalpark in Botswana.
(Mit freundlicher
Genehmigung von Brian Walker.)
Im
Jahr 1982 trocknete er erneut aus und ist bis heute in diesem Zustand
geblieben. Kurz nachdem der Kanal in den ersten Jahren des 20. Jahrhunderts
kein Wasser mehr führte, gab es in der Gegend eine große Rinderpestepidemie.
Diese beiden Ereignisse sorgten gemeinsam für die Entstehung der heutigen
Akazienwälder, und das ging folgendermaßen: Wegen des Wassermangels suchten
die Elefanten sich in anderen Gegenden etwas zu trinken (und außerdem wurde
ihre Zahl durch Jäger dezimiert). Und die Rinderpest vernichtete weitgehend
die Population der Huftiere. Die Folge war, daß es in der Region kaum noch
grasende Tiere gab, so daß die Akazienschößlinge (eine Lieblingsspeise
vieler Pflanzenfresser) zu Bäumen heranwachsen konnten. Als Elefanten und
Huftiere zurückkehrten, war die Gegend von großen Akazienwäldern bedeckt.
»Das heute zu beobachtende Nebeneinander von vielen Elefanten und
umfangreichen Akaziengehölzen ist zeitlich sehr eng begrenzt und offenbar nicht
nachhaltig«, meint Brian Walker, der die Region eingehend untersucht hat.12
Es
ist nicht nachhaltig, weil Akazienschößlinge nicht bis zum Erwachsenenalter
überleben, solange es in der Gegend gesunde Elefanten- und Huftierpopulationen gibt. Wenn die Wälder wieder gedeihen sollten, müßte
man die Tiere fernhalten. Walker stellt fest: »Die Frage ist, ob Verwalter
und Touristen bereit sind, eine Phase von zehn bis 15 Jahren zu akzeptieren,
in der praktisch keine Tiere zu sehen sind.« Vermutlich sind sie das nicht.
Die derzeitige Artenvielfalt in dem Park ist zwar etwas Natürliches, aber sie
hat ihre Ursache in tiefgreifenden Umweltveränderungen, die sich im Laufe
vieler Jahrzehnte abgespielt haben. Und Parkverwalter haben häufig etwas
gegen Veränderungen, vor allem wenn man etwas Wertvolles verschwinden sieht.
Ökosysteme sind sowohl räumlich als auch zeitlich in ständiger Umwälzung
begriffen, und immer werden manche Populationen abnehmen, während andere
gedeihen und sich vermehren. Der ständige Wandel ist als Motor der
Artenvielfalt unentbehrlich.
»Die Naturschützer sollten sich weniger
Gedanken über die dauerhafte Erhaltung einzelner Tier- und Pflanzenarten
machen«, warnt Walker, »und statt dessen lieber überlegen, wie man die
Natur und die Vielfalt der ökologischen Prozesse als Ganzes erhalten kann.«13)
Vor
dem Hintergrund dessen, was über das Wesen der Ökosysteme gesagt wurde, und
mit dem Wissen um Chaos und den dynamischen Aufbau der Lebensgemeinschaften
können wir erkennen, daß Walkers Mahnung begründet ist. Aber wie bei allen
Angelegenheiten der Menschen ist es schwierig, etwas in Gang zu setzen, das
viele Jahrzehnte dauert. Niemand möchte untätig herumsitzen und zusehen, wie
Wälder schrumpfen oder wie Tiere verhungern oder verdursten. Aber genau das
müssen wir letztlich wahrscheinlich tun.
208
#
www.detopia.de
^^^^